Menštruačný cyklus, jeho regulácia. Neurohumorálna regulácia ženskej reprodukčnej funkcie
Kapitola 2. Neuroendokrinná regulácia menštruačného cyklu
Kapitola 2. Neuroendokrinná regulácia menštruačného cyklu
Menštruačný cyklus - geneticky podmienené, cyklicky sa opakujúce zmeny v organizme ženy, najmä v častiach reprodukčného systému, ktorých klinickým prejavom je výtok krvi z pohlavného traktu (menštruácia).
Menštruačný cyklus vzniká po menarche (prvá menštruácia) a pretrváva počas celého reprodukčného (plodného) obdobia života ženy až do menopauzy (posledná menštruácia). Cyklické zmeny v tele ženy sú zamerané na možnosť reprodukcie potomstva a sú dvojfázového charakteru: 1. (folikulárna) fáza cyklu je určená rastom a dozrievaním folikulu a vajíčka vo vaječníku, po ktorom folikul praskne a vajíčko ho opustí - ovulácia; 2. (luteálna) fáza je spojená s tvorbou žltého telieska. Súčasne v cyklickom režime dochádza k postupným zmenám v endometriu: regenerácia a proliferácia funkčnej vrstvy, po ktorej nasleduje sekrečná transformácia žliaz. Zmeny v endometriu končia deskvamáciou funkčnej vrstvy (menštruácia).
Biologický význam zmien, ku ktorým dochádza počas menštruačného cyklu vo vaječníkoch a endometriu, spočíva v zabezpečení reprodukčnej funkcie po dozretí vajíčka, jeho oplodnení a uhniezdení embrya v maternici. Ak nedôjde k oplodneniu vajíčka, funkčná vrstva endometria sa odmietne, z genitálneho traktu sa objavia sekréty krvi a v reprodukčnom systéme sa znova a v rovnakom poradí vyskytujú procesy zamerané na zabezpečenie dozrievania vajíčka.
menštruácia - ide o výtok krvi z pohlavného traktu, ktorý sa opakuje v určitých intervaloch počas celého reprodukčného obdobia, s výnimkou tehotenstva a laktácie. Menštruácia začína na konci luteálnej fázy menštruačného cyklu v dôsledku odlupovania funkčnej vrstvy endometria. Prvá menštruácia (menarhe) vyskytuje sa vo veku 10-12 rokov. Najbližšieho 1-1,5 roka môže byť menštruácia nepravidelná a až potom sa vytvorí pravidelný menštruačný cyklus.
Prvý deň menštruácie sa podmienečne považuje za 1. deň menštruačného cyklu a trvanie cyklu sa vypočíta ako interval medzi prvými dňami dvoch po sebe nasledujúcich menštruácií.
Vonkajšie parametre normálneho menštruačného cyklu:
Trvanie - od 21 do 35 dní (60% žien má priemernú dĺžku cyklu 28 dní);
Trvanie menštruačného toku je od 3 do 7 dní;
Množstvo straty krvi v dňoch menštruácie je 40-60 ml (v priemere
50 ml).
Procesy, ktoré zabezpečujú normálny priebeh menštruačného cyklu, sú regulované jediným funkčne prepojeným neuroendokrinným systémom vrátane centrálnych (integrujúcich) oddelení, periférnych (efektorových) štruktúr, ako aj medzičlánkov.
Fungovanie reprodukčného systému je zabezpečené striktne geneticky naprogramovanou interakciou piatich hlavných úrovní, z ktorých každá je regulovaná nadložnými štruktúrami podľa princípu priamych a inverzných, pozitívnych a negatívnych vzťahov (obr. 2.1).
Prvá (najvyššia) úroveň regulácie reprodukčný systém sú kôra a extrahypotalamických cerebrálnych štruktúr
(limbický systém, hipokampus, amygdala). Primeraný stav centrálneho nervového systému zabezpečuje normálne fungovanie všetkých základných častí reprodukčného systému. Rôzne organické a funkčné zmeny v kôre a podkôrových štruktúrach môžu viesť k nepravidelnostiam menštruačného cyklu. Možnosť zastavenia menštruácie pri silnom strese (strata blízkych, vojnové podmienky a pod.) alebo bez zjavných vonkajších vplyvov s celkovou psychickou nerovnováhou („falošné tehotenstvo“ je oneskorenie menštruácie so silnou túžbou po otehotnení alebo naopak s jeho strach) je dobre známy. ).
Špecifické neuróny mozgu dostávajú informácie o stave vonkajšieho aj vnútorného prostredia. Vnútorná expozícia sa uskutočňuje pomocou špecifických receptorov pre ovariálne steroidné hormóny (estrogény, progesterón, androgény), ktoré sa nachádzajú v centrálnom nervovom systéme. V reakcii na vplyv faktorov prostredia na mozgovú kôru a extrahypotalamické štruktúry dochádza k syntéze, vylučovaniu a metabolizmu. neurotransmitery a neuropeptidy. Neurotransmitery a neuropeptidy zase ovplyvňujú syntézu a uvoľňovanie hormónov neurosekrečnými jadrami hypotalamu.
K tomu najdôležitejšiemu neurotransmitery, tie. Látky-prenášače nervových impulzov zahŕňajú norepinefrín, dopamín, kyselinu γ-aminomaslovú (GABA), acetylcholín, serotonín a melatonín. Norepinefrín, acetylcholín a GABA stimulujú uvoľňovanie gonadotropného uvoľňujúceho hormónu (GnRH) hypotalamom. Dopamín a serotonín znižujú frekvenciu a amplitúdu produkcie GnRH počas menštruačného cyklu.
Neuropeptidy(endogénne opioidné peptidy, neuropeptid Y, galanín) sa podieľajú aj na regulácii funkcie reprodukčného systému. Opioidné peptidy (endorfíny, enkefalíny, dynorfíny), viažuce sa na opiátové receptory, vedú k potlačeniu syntézy GnRH v hypotalame.
Ryža. 2.1. Hormonálna regulácia v systéme hypotalamus - hypofýza - periférne endokrinné žľazy - cieľové orgány (schéma): RG - uvoľňujúce hormóny; TSH - hormón stimulujúci štítnu žľazu; ACTH - adrenooktotropný hormón; FSH - folikuly stimulujúci hormón; LH - luteinizačný hormón; Prl - prolaktín; P - progesterón; E - estrogény; A - androgény; P - relaxín; I - ingi-bin; T 4 - tyroxín, ADH - antidiuretický hormón (vazopresín)
Druhá úroveň regulácia reprodukčnej funkcie je hypotalamus. Napriek svojej malej veľkosti sa hypotalamus podieľa na regulácii sexuálneho správania, riadi vegetovaskulárne reakcie, telesnú teplotu a ďalšie životne dôležité funkcie tela.
Hypofyziotropná zóna hypotalamu reprezentované skupinami neurónov, ktoré tvoria neurosekrečné jadrá: ventromediálne, dorzomediálne, oblúkové, supraoptické, paraventrikulárne. Tieto bunky majú vlastnosti neurónov (reprodukujúcich elektrické impulzy) a endokrinných buniek, ktoré produkujú špecifické neurosekréty s diametrálne opačnými účinkami (liberíny a statíny). liberíny, alebo uvoľňujúce faktory, stimulovať uvoľňovanie vhodných tropických hormónov v prednej hypofýze. statíny majú inhibičný účinok na ich uvoľňovanie. V súčasnosti je známych sedem liberínov, ktoré sú svojou podstatou dekapeptidy: tyreoliberín, kortikoliberín, somatoliberín, melanoliberín, foliberín, luliberín, prolaktoliberín, ako aj tri statíny: melanostatín, somatostatín, prolaktostatín alebo faktor inhibujúci prolaktín.
Luliberín alebo hormón uvoľňujúci luteinizačný hormón (LHRH) bol izolovaný, syntetizovaný a podrobne opísaný. Doteraz nebolo možné izolovať a syntetizovať folikuly stimulujúci hormón. Zistilo sa však, že RGHL a jeho syntetické analógy stimulujú uvoľňovanie nielen LH, ale aj FSH gonadotropmi. V tejto súvislosti sa pre gonadotropné liberíny ujal jeden termín – „homón uvoľňujúci gonadotropín“ (GnRH), ktorý je v skutočnosti synonymom pre luliberín (RHRH).
Hlavným miestom sekrécie GnRH sú oblúkovité, supraoptické a paraventrikulárne jadrá hypotalamu. Oblúkové jadrá reprodukujú sekrečný signál s frekvenciou približne 1 pulz za 1-3 hodiny, t.j. v pulzujúca alebo cirhorálny režim (kruhový- okolo hodiny). Tieto impulzy majú určitú amplitúdu a spôsobujú periodický tok GnRH cez portálny krvný obeh do buniek adenohypofýzy. V závislosti od frekvencie a amplitúdy pulzov GnRH adenohypofýza prevažne vylučuje LH alebo FSH, čo zase spôsobuje morfologické a sekrečné zmeny vo vaječníkoch.
Hypotalamo-hypofyzárna oblasť má špeciálnu cievnu sieť tzv portálový systém. Charakteristickým rysom tejto vaskulárnej siete je schopnosť prenášať informácie z hypotalamu do hypofýzy a naopak (z hypofýzy do hypotalamu).
Regulácia uvoľňovania prolaktínu je do značnej miery pod vplyvom statínov. Dopamín, produkovaný v hypotalame, inhibuje uvoľňovanie prolaktínu z laktotrofov adenohypofýzy. Tyreoliberín, ako aj serotonín a endogénne opioidné peptidy prispievajú k zvýšeniu sekrécie prolaktínu.
Okrem liberínov a statínov sa v hypotalame (supraoptické a paraventrikulárne jadrá) tvoria dva hormóny: oxytocín a vazopresín (antidiuretický hormón). Granule obsahujúce tieto hormóny migrujú z hypotalamu pozdĺž axónov veľkých bunkových neurónov a hromadia sa v zadnej hypofýze (neurohypofýze).
Tretia úroveň regulácia reprodukčnej funkcie je hypofýza, pozostáva z predného, zadného a stredného (stredného) laloku. Priamo súvisí s reguláciou reprodukčnej funkcie predný lalok (adenohypofýza) . Vplyvom hypotalamu sa v adenohypofýze vylučujú gonadotropné hormóny – FSH (alebo folitropín), LH (alebo lutropín), prolaktín (Prl), ACTH, somatotropný (STH) a hormón stimulujúci štítnu žľazu (TSH). Normálne fungovanie reprodukčného systému je možné len pri vyváženom výbere každého z nich.
Gonadotropné hormóny (FSH, LH) prednej hypofýzy sú pod kontrolou GnRH, ktorý stimuluje ich sekréciu a uvoľňovanie do krvného obehu. Pulzujúci charakter sekrécie FSH, LH je výsledkom "priamych signálov" z hypotalamu. Frekvencia a amplitúda impulzov sekrécie GnRH sa mení v závislosti od fáz menštruačného cyklu a ovplyvňuje koncentráciu a pomer FSH/LH v krvnej plazme.
FSH stimuluje rast folikulov vo vaječníku a dozrievanie vajíčka, proliferáciu granulózových buniek, tvorbu FSH a LH receptorov na povrchu granulózových buniek, aktivitu aromatázy v dozrievacom folikule (to zvyšuje konverziu androgény na estrogény), produkciu inhibínu, aktivínu a inzulínu podobných rastových faktorov.
LH podporuje tvorbu androgénov v bunkách theca, zabezpečuje ovuláciu (spolu s FSH), stimuluje syntézu progesterónu v luteinizovaných granulózových bunkách (žlté teliesko) po ovulácii.
Prolaktín má rôzne účinky na telo ženy. Jeho hlavnou biologickou úlohou je stimulovať rast mliečnych žliaz, regulovať laktáciu; má tiež tuk mobilizujúci a hypotenzívny účinok, riadi sekréciu progesterónu žltým telieskom tým, že v ňom aktivuje tvorbu LH receptorov. Počas tehotenstva a dojčenia sa hladina prolaktínu v krvi zvyšuje. Hyperprolaktinémia vedie k narušeniu rastu a dozrievania folikulov vo vaječníku (anovulácia).
Zadná hypofýza (neurohypofýza) nie je žľaza s vnútornou sekréciou, ale iba ukladá hormóny hypotalamu (oxytocín a vazopresín), ktoré sú v tele vo forme proteínového komplexu.
vaječníkov vzťahovať do štvrtej úrovne reguláciu reprodukčného systému a vykonávajú dve hlavné funkcie. Vo vaječníkoch dochádza k cyklickému rastu a dozrievaniu folikulov, dozrievaniu vajíčka, t.j. vykonáva sa generatívna funkcia, ako aj syntéza pohlavných steroidov (estrogény, androgény, progesterón) - hormonálna funkcia.
Hlavnou morfofunkčnou jednotkou vaječníka je folikul. Pri narodení obsahujú vaječníky dievčaťa približne 2 milióny primordiálnych folikulov. Väčšina z nich (99 %) prejde počas života atréziou (spätným vývojom folikulov). Len veľmi malá časť z nich (300 – 400) prejde úplným vývojovým cyklom – od prvotného po predovulačný s následným vytvorením žltého telieska. V čase menarche obsahujú vaječníky 200-400 tisíc prvotných folikulov.
Ovariálny cyklus pozostáva z dvoch fáz: folikulárnej a luteálnej. Folikulárna fáza začína po menštruácii, spojená s rastom
a dozrievanie folikulov a končí ovuláciou. luteálna fáza zaberá interval po ovulácii až do začiatku menštruácie a je spojený s tvorbou, vývojom a regresiou žltého telieska, ktorého bunky vylučujú progesterón.
V závislosti od stupňa zrelosti sa rozlišujú štyri typy folikulov: primordiálne, primárne (preantrálne), sekundárne (antrálne) a zrelé (preovulačné, dominantné) (obr. 2.2).

Ryža. 2.2.Štruktúra vaječníka (diagram). Etapy vývoja dominantného folikulu a žltého tela: 1 - väzivo vaječníka; 2 - proteínový plášť; 3 - cievy vaječníka (konečná vetva ovariálnej tepny a žily); 4 - primordiálny folikul; 5 - preantrálny folikul; 6 - antrálny folikul; 7 - predovulačný folikul; 8 - ovulácia; 9 - corpus luteum; 10 - biele telo; 11 - vajíčko (oocyt); 12 - bazálna membrána; 13 - folikulárna tekutina; 14 - vaječný tuberkul; 15 - škrupina theca; 16 - lesklá škrupina; 17 - granulózne bunky
Primordiálny folikul pozostáva z nezrelého vajíčka (oocytu) v profáze 2. meiotického delenia, ktoré je obklopené jednou vrstvou granulóznych buniek.
AT preantrálny (primárny) folikul oocyt sa zväčšuje. Bunky zrnitého epitelu proliferujú a zaokrúhľujú sa, čím vytvárajú zrnitú vrstvu folikulu. Z okolitej strómy sa vytvára spojivovo-netkaná pošva – theka (theca).
Antrálny (sekundárny) folikul charakterizovaný ďalším rastom: pokračuje proliferácia buniek granulóznej vrstvy, ktoré produkujú folikulárnu tekutinu. Výsledná tekutina vytlačí vajíčko na perifériu, kde bunky zrnitej vrstvy vytvoria vajíčkovú tuberkulózu (cumulus oophorus). Membrána spojivového tkaniva folikulu je jasne rozlíšená na vonkajšiu a vnútornú. Vnútorná škrupina (the-ca interna) pozostáva z 2-4 vrstiev buniek. vonkajšia škrupina (theca externa) sa nachádza nad vnútornou a je reprezentovaná diferencovanou strómou spojivového tkaniva.
AT predovulačný (dominantný) folikul vajíčko umiestnené na tuberkulu vajíčka je pokryté membránou nazývanou zona pellucida (zona pellucida). V oocyte dominantného folikulu sa obnoví proces meiózy. Počas dozrievania dochádza v preovulačnom folikule k stonásobnému zväčšeniu objemu folikulárnej tekutiny (priemer folikulu dosahuje 20 mm) (obr. 2.3).
Počas každého menštruačného cyklu začne rásť 3 až 30 primordiálnych folikulov, ktoré sa transformujú na preantrálne (primárne) folikuly. V nasledujúcom menštruačnom cykle pokračuje folikulo-logogenéza a len jeden folikul sa vyvíja od preantrálneho po preovulačný. Počas rastu folikulu z preantrálneho do antrálneho

Ryža. 2.3. Dominantný folikul vo vaječníku. Laparoskopia
granulózne bunky syntetizujú anti-mullerovský hormón, ktorý prispieva k jeho rozvoju. Zostávajúce folikuly, ktoré pôvodne vstúpili do rastu, podliehajú atrézii (degenerácii).
ovulácia - prasknutie preovulačného (dominantného) folikulu a uvoľnenie vajíčka z neho do brušnej dutiny. Ovuláciu sprevádza krvácanie zo zničených kapilár obklopujúcich bunky théky (obr. 2.4).
Po uvoľnení vajíčka výsledné kapiláry rýchlo rastú do zostávajúcej dutiny folikulu. Bunky granulózy prechádzajú luteinizáciou, ktorá sa morfologicky prejavuje zväčšením ich objemu a tvorbou lipidových inklúzií – a corpus luteum(obr. 2.5).

Ryža. 2.4. Ovariálny folikul po ovulácii. Laparoskopia

Ryža. 2.5.Žlté teliesko vaječníka. Laparoskopia
Žlté telo - prechodná hormonálne aktívna formácia, fungujúca 14 dní, bez ohľadu na celkové trvanie menštruačného cyklu. Ak nedôjde k otehotneniu, žlté teliesko ustúpi, ak však dôjde k oplodneniu, funguje až do vytvorenia placenty (12. týždeň tehotenstva).
Hormonálna funkcia vaječníkov
Rast, dozrievanie folikulov vo vaječníkoch a tvorba žltého telieska sú sprevádzané produkciou pohlavných hormónov tak granulóznymi bunkami folikulu, ako aj bunkami vnútornej teky a v menšej miere aj vonkajšej teky. Pohlavné steroidné hormóny zahŕňajú estrogény, progesterón a androgény. Východiskovým materiálom pre tvorbu všetkých steroidných hormónov je cholesterol. Až 90 % steroidných hormónov je vo viazanom stave a len 10 % neviazaných hormónov má svoj biologický účinok.
Estrogény sú rozdelené do troch frakcií s rôznou aktivitou: estradiol, estriol, estrón. Estrón - najmenej aktívna frakcia, je vylučovaná vaječníkmi hlavne počas starnutia - v postmenopauze; najaktívnejšou frakciou je estradiol, je významný pri nástupe a udržiavaní tehotenstva.
Množstvo pohlavných hormónov sa počas menštruačného cyklu mení. S rastom folikulu sa zvyšuje syntéza všetkých pohlavných hormónov, ale hlavne estrogénu. V období po ovulácii a pred nástupom menštruácie sa progesterón syntetizuje prevažne vo vaječníkoch, vylučovaný bunkami žltého telieska.
Androgény (androstendión a testosterón) sú produkované tekálnymi bunkami folikulu a intersticiálnymi bunkami. Ich hladina počas menštruačného cyklu sa nemení. Androgény, ktoré sa dostanú do buniek granulózy, aktívne podstupujú aromatizáciu, čo vedie k ich premene na estrogény.
Okrem steroidných hormónov vylučujú vaječníky aj ďalšie biologicky aktívne zlúčeniny: prostaglandíny, oxytocín, vazopresín, relaxín, epidermálny rastový faktor (EGF), inzulínu podobné rastové faktory (IPFR-1 a IPFR-2). Predpokladá sa, že rastové faktory prispievajú k proliferácii granulóznych buniek, rastu a dozrievaniu folikulu a k selekcii dominantného folikulu.
V procese ovulácie zohrávajú určitú úlohu prostaglandíny (F 2a a E 2), ako aj proteolytické enzýmy obsiahnuté vo folikulárnej tekutine, kolagenáza, oxytocín, relaxín.
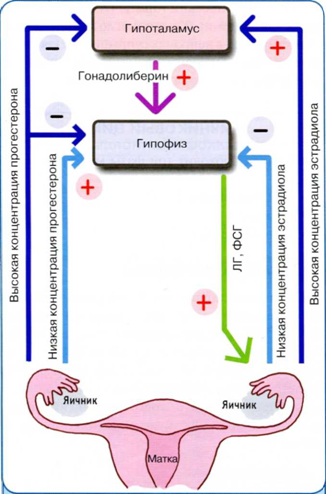
Cyklická aktivita reprodukčného systému je determinovaná princípmi priamej a spätnej väzby, ktorú zabezpečujú špecifické hormonálne receptory v každej z väzieb. Priamou väzbou je stimulačný účinok hypotalamu na hypofýzu a následná tvorba pohlavných steroidov vo vaječníku. Spätná väzba je určená vplyvom zvýšenej koncentrácie pohlavných steroidov na nadložné hladiny, blokujúc ich aktivitu.
Pri interakcii väzieb reprodukčného systému sa rozlišujú "dlhé", "krátke" a "ultrakrátke" slučky. "Dlhá" slučka - vplyv cez receptory hypotalamo-hypofyzárneho systému na produkciu pohlavných hormónov. "Krátka" slučka určuje spojenie medzi hypofýzou a hypotalamom, "ultrashort" slučka určuje spojenie medzi hypotalamom a nervovými bunkami, ktoré pod vplyvom elektrických stimulov vykonávajú lokálnu reguláciu pomocou neurotransmiterov, neuropeptidy a neuromodulátory.
Folikulárna fáza
Pulzujúca sekrécia a uvoľňovanie GnRH vedie k uvoľňovaniu FSH a LH z prednej hypofýzy. LH podporuje syntézu androgénov bunkami theca vo folikule. FSH pôsobí na vaječníky a vedie k rastu folikulov a dozrievaniu oocytov. Súčasne zvyšujúca sa hladina FSH stimuluje produkciu estrogénov v granulózových bunkách aromatizáciou androgénov vytvorených v tekálnych bunkách folikulu a tiež podporuje sekréciu inhibínu a IPFR-1-2. Pred ovuláciou sa zvyšuje počet receptorov pre FSH a LH v bunkách théky a granulózy (obr. 2.6).
Ovulácia nastáva uprostred menštruačného cyklu, 12-24 hodín po dosiahnutí vrcholu estradiolu, čo spôsobuje zvýšenie frekvencie a amplitúdy sekrécie GnRH a prudký predovulačný vzostup sekrécie LH typom „pozitívnej spätnej väzby“. Na tomto pozadí sa aktivujú proteolytické enzýmy – kolagenáza a plazmín, ktoré ničia kolagén steny folikulu a tým znižujú jeho pevnosť. Zároveň pozorované zvýšenie koncentrácie prostaglandínu F 2a, ako aj oxytocínu, vyvoláva ruptúru folikulu v dôsledku ich stimulácie kontrakcie hladkého svalstva a vypudenia oocytu s vajcovodným tuberkulom z dutiny folikul. Prasknutie folikulu je tiež uľahčené zvýšením koncentrácie prostaglandínu E2 a relaxínu v ňom, ktoré znižujú tuhosť jeho stien.
luteálna fáza
Po ovulácii hladina LH klesá v pomere k „ovulačnému vrcholu“. Toto množstvo LH však stimuluje proces luteinizácie granulóznych buniek zostávajúcich vo folikule, ako aj prevládajúcu sekréciu progesterónu vytvoreným žltým telieskom. K maximálnej sekrécii progesterónu dochádza 6. – 8. deň existencie žltého telieska, čo zodpovedá 20. – 22. dňu menštruačného cyklu. Postupne do 28. – 30. dňa menštruačného cyklu klesá hladina progesterónu, estrogénu, LH a FSH, dochádza k regresii žltého telieska a nahrádza ho väzivo (biele teliesko).
Piata úroveň regulácie reprodukčných funkcií sú cieľové orgány citlivé na kolísanie hladiny pohlavných steroidov: maternica, vajíčkovody, pošvová sliznica, ale aj mliečne žľazy, vlasové folikuly, kosti, tukové tkanivo, centrálny nervový systém.
Ovariálne steroidné hormóny ovplyvňujú metabolické procesy v orgánoch a tkanivách, ktoré majú špecifické receptory. Tieto receptory môžu byť

Ryža. 2.6. Hormonálna regulácia menštruačného cyklu (schéma): a - zmeny hladiny hormónov; b - zmeny vo vaječníku; c - zmeny v endometriu
cytoplazmatické aj jadrové. Cytoplazmatické receptory sú vysoko špecifické pre estrogén, progesterón a testosterón. Steroidy prenikajú do cieľových buniek väzbou na špecifické receptory – respektíve na estrogén, progesterón, testosterón. Výsledný komplex vstupuje do bunkového jadra, kde spojením s chromatínom zabezpečuje syntézu špecifických tkanivových proteínov prostredníctvom transkripcie messenger RNA.
Uterus pozostáva z vonkajšieho (serózneho) krytu, myometria a endometria. Endometrium morfologicky pozostáva z dvoch vrstiev: bazálnej a funkčnej. Bazálna vrstva sa počas menštruačného cyklu výrazne nemení. Funkčná vrstva endometria prechádza štrukturálnymi a morfologickými zmenami, ktoré sa prejavujú postupnou zmenou štádií proliferácia, sekrécia, deskvamácia nasledovaný
regenerácia. Cyklická sekrécia pohlavných hormónov (estrogénov, progesterónu) vedie k dvojfázovým zmenám v endometriu, ktorých cieľom je vnímanie oplodneného vajíčka.
Cyklické zmeny v endometriu sa týkajú jeho funkčnej (povrchovej) vrstvy, pozostávajúcej z kompaktných epiteliálnych buniek, ktoré sú počas menštruácie odmietnuté. Bazálna vrstva, ktorá sa v tomto období neodmieta, zabezpečuje obnovu funkčnej vrstvy.
V endometriu sa počas menštruačného cyklu vyskytujú tieto zmeny: deskvamácia a odmietnutie funkčnej vrstvy, regenerácia, fáza proliferácie a fáza sekrécie.
Transformácia endometria nastáva pod vplyvom steroidných hormónov: fáza proliferácie - pod prevažujúcim pôsobením estrogénov, fáza sekrécie - pod vplyvom progesterónu a estrogénov.
Proliferačná fáza(zodpovedá folikulárnej fáze vo vaječníkoch) trvá v priemere 12-14 dní, počnúc 5. dňom cyklu. V tomto období sa vytvára nová povrchová vrstva s predĺženými tubulárnymi žľazami lemovanými cylindrickým epitelom so zvýšenou mitotickou aktivitou. Hrúbka funkčnej vrstvy endometria je 8 mm (obr. 2.7).
Fáza sekrécie (luteálna fáza vo vaječníkoch) spojené s činnosťou žltého telieska, trvá 14±1 deň. V tomto období začína epitel endometriálnych žliaz produkovať sekrét obsahujúci kyslé glykozaminoglykány, glykoproteíny, glykogén (obr. 2.8).

Ryža. 2.7. Endometrium vo fáze proliferácie (stredné štádium). Farbené hematoxylínom a eozínom, × 200. Foto O.V. Zayratyan

Ryža. 2.8. Endometrium vo fáze sekrécie (stredné štádium). Zafarbené hematoxylínom a eozínom, ×200. Foto O.V. Zayratyan
Aktivita sekrécie je najvyššia v 20.-21. deň menštruačného cyklu. Do tejto doby sa maximálne množstvo proteolytických enzýmov nachádza v endometriu a deciduálne transformácie sa vyskytujú v stróme. Dochádza k prudkej vaskularizácii strómy – špirálovité tepny funkčnej vrstvy sú kľukaté, tvoria „spletence“, žily sú rozšírené. Takéto zmeny v endometriu, pozorované na 20. až 22. deň (6. až 8. deň po ovulácii) 28-dňového menštruačného cyklu, poskytujú najlepšie podmienky pre implantáciu oplodneného vajíčka.
Na 24-27 deň je v dôsledku začiatku regresie žltého telieska a zníženia koncentrácie ním produkovaného progesterónu narušený endometriálny trofizmus a postupne sa v ňom zvyšujú degeneratívne zmeny. Z granulárnych buniek endometriálnej strómy sa uvoľňujú granuly obsahujúce relaxín, ktorý pripravuje menštruačné odmietnutie sliznice. V povrchových oblastiach kompaktnej vrstvy je zaznamenaná lacunárna expanzia kapilár a krvácanie v stróme, ktoré je možné zistiť 1 deň pred začiatkom menštruácie.
Menštruácia zahŕňa deskvamáciu, odmietnutie a regeneráciu funkčnej vrstvy endometria. V dôsledku regresie žltého telieska a prudkého poklesu obsahu pohlavných steroidov v endometriu sa zvyšuje hypoxia. Nástup menštruácie je uľahčený predĺženým kŕčom tepien, čo vedie k stagnácii krvi a tvorbe krvných zrazenín. Tkanivovú hypoxiu (tkanivovú acidózu) zhoršuje zvýšená permeabilita endotelu, krehkosť cievnych stien, početné malé krvácania a masívna leukémia.
cytickej infiltrácie. Lysozomálne proteolytické enzýmy uvoľňované z leukocytov zvyšujú topenie tkanivových prvkov. Po dlhotrvajúcom spazme ciev dochádza k ich paretickej expanzii so zvýšeným prietokom krvi. Súčasne dochádza k zvýšeniu hydrostatického tlaku v mikrovaskulatúre a k prasknutiu stien ciev, ktoré do tejto doby do značnej miery stratili svoju mechanickú pevnosť. Na tomto pozadí dochádza k aktívnej deskvamácii nekrotických oblastí funkčnej vrstvy endometria. Do konca 1. dňa menštruácie sú odmietnuté 2/3 funkčnej vrstvy a jej úplná deskvamácia sa zvyčajne končí na 3. deň menštruačného cyklu.
Regenerácia endometria začína ihneď po odmietnutí nekrotickej funkčnej vrstvy. Základom pre regeneráciu sú epitelové bunky strómy bazálnej vrstvy. Za fyziologických podmienok už na 4. deň cyklu dochádza k epitelizácii celého povrchu rany na sliznici. Potom opäť nasledujú cyklické zmeny v endometriu – fázy proliferácie a sekrécie.
Postupné zmeny v endometriu počas cyklu – proliferácia, sekrécia a menštruácia – závisia nielen od cyklických výkyvov hladiny pohlavných steroidov v krvi, ale aj od stavu tkanivových receptorov pre tieto hormóny.
Koncentrácia jadrových estradiolových receptorov sa zvyšuje až do polovice cyklu, pričom vrchol dosahuje v neskoršom období fázy proliferácie endometria. Po ovulácii dochádza k rýchlemu poklesu koncentrácie jadrových estradiolových receptorov, ktorý pokračuje až do neskorej sekrečnej fázy, kedy je ich expresia výrazne nižšia ako na začiatku cyklu.
Funkčný stav vajíčkovodov sa líši v závislosti od fázy menštruačného cyklu. Takže v luteálnej fáze cyklu sa aktivuje ciliárny aparát ciliárneho epitelu a kontraktilná aktivita svalovej vrstvy zameraná na optimálny transport pohlavných gamét do dutiny maternice.
Zmeny v extragenitálnych cieľových orgánoch
Všetky pohlavné hormóny nielen určujú funkčné zmeny v samotnom reprodukčnom systéme, ale tiež aktívne ovplyvňujú metabolické procesy v iných orgánoch a tkanivách, ktoré majú receptory pre pohlavné steroidy.
V pokožke sa vplyvom estradiolu a testosterónu aktivuje syntéza kolagénu, ktorá pomáha udržiavať jej elasticitu. Zvýšený kožný maz, akné, folikulitída, pórovitosť kože a nadmerné ochlpenie sa vyskytujú so zvýšením hladiny androgénov.
V kostiach estrogény, progesterón a androgény podporujú normálnu prestavbu tým, že bránia kostnej resorpcii. Rovnováha pohlavných steroidov ovplyvňuje metabolizmus a distribúciu tukového tkaniva v ženskom tele.
Účinok pohlavných hormónov na receptory v centrálnom nervovom systéme a hipokampálnych štruktúrach je spojený so zmenami v emocionálnej sfére a
reakcie u ženy v dňoch pred menštruáciou - fenomén "menštruačnej vlny". Tento jav sa prejavuje nerovnováhou v procesoch aktivácie a inhibície v mozgovej kôre, kolísaním sympatikového a parasympatického nervového systému (postihuje najmä kardiovaskulárny systém). Vonkajšími prejavmi týchto výkyvov sú zmeny nálady a podráždenosť. U zdravých žien tieto zmeny neprekračujú fyziologické hranice.
Vplyv štítnej žľazy a nadobličiek na reprodukčnú funkciu
Štítna žľaza produkuje dva hormóny kyseliny jódamínovej - trijódtyronín (T 3) a tyroxín (T 4), ktoré sú najdôležitejšími regulátormi metabolizmu, vývoja a diferenciácie všetkých telesných tkanív, najmä tyroxínu. Hormóny štítnej žľazy majú určitý vplyv na proteín-syntetickú funkciu pečene, stimulujú tvorbu globulínu, ktorý viaže pohlavné steroidy. To sa odráža v rovnováhe voľných (aktívnych) a viazaných ovariálnych steroidov (estrogény, androgény).
Pri nedostatku T 3 a T 4 sa zvyšuje sekrécia tyreoliberínu, ktorý aktivuje nielen tyreotrofy, ale aj laktotrofy hypofýzy, čo často spôsobuje hyperprolaktinémiu. Paralelne sa znižuje sekrécia LH a FSH s inhibíciou folikulov a steroidogenézy vo vaječníkoch.
Zvýšenie hladiny T3 a T4 je sprevádzané výrazným zvýšením koncentrácie globulínu, ktorý viaže pohlavné hormóny v pečeni a vedie k zníženiu voľnej frakcie estrogénov. Hypoestrogenizmus zase vedie k porušeniu dozrievania folikulov.
Nadobličky. Normálne je produkcia androgénov – androstendiónu a testosterónu – v nadobličkách rovnaká ako vo vaječníkoch. V nadobličkách dochádza k tvorbe DHEA a DHEA-S, pričom tieto androgény sa vo vaječníkoch prakticky nesyntetizujú. DHEA-S, ktorý sa vylučuje v najväčšom množstve (v porovnaní s inými adrenálnymi androgénmi), má relatívne nízku androgénnu aktivitu a slúži ako akási rezervná forma androgénov. Suprarenálne androgény spolu s androgénmi ovariálneho pôvodu sú substrátom pre extragonadálnu produkciu estrogénu.
Posúdenie stavu reprodukčného systému podľa testov funkčnej diagnostiky
Už dlhé roky sa v gynekologickej praxi používajú takzvané testy funkčnej diagnostiky stavu reprodukčného systému. Hodnota týchto pomerne jednoduchých štúdií sa zachovala až do súčasnosti. Najpoužívanejšie je meranie bazálnej teploty, hodnotenie fenoménu „zrenice“ a stavu hlienu krčka maternice (jeho kryštalizácia, rozťažnosť), ako aj výpočet karyopyknotického indexu (KPI,%) vaginálnej epitelu (obr. 2.9).

Ryža. 2.9. Funkčné diagnostické testy pre dvojfázový menštruačný cyklus
Test bazálnej teploty je založená na schopnosti progesterónu (vo zvýšenej koncentrácii) priamo ovplyvňovať termoregulačné centrum v hypotalame. Pod vplyvom progesterónu v 2. (luteálnej-novej) fáze menštruačného cyklu dochádza k prechodnej hypertermickej reakcii.
Pacient denne meria teplotu v konečníku ráno bez toho, aby vstal z postele. Výsledky sú zobrazené graficky. Pri bežnom dvojfázovom menštruačnom cykle nepresiahne bazálna teplota v 1. (folikulárnej) fáze menštruačného cyklu 37 °C, v 2. (luteálnej) fáze dochádza k zvýšeniu rektálnej teploty o 0,4-0,8 °C. v porovnaní s počiatočnou hodnotou. V deň menštruácie alebo 1 deň pred jej začiatkom dochádza k regresii žltého telieska vo vaječníku, k zníženiu hladiny progesterónu, a teda k zníženiu bazálnej teploty na pôvodné hodnoty.
Pretrvávajúci dvojfázový cyklus (bazálna teplota by sa mala merať počas 2-3 menštruačných cyklov) naznačuje, že nastala ovulácia a funkčná užitočnosť žltého telieska. Neprítomnosť zvýšenia teploty v 2. fáze cyklu naznačuje absenciu ovulácie (anovulácie); oneskorenie vzostupu, jeho krátke trvanie (zvýšenie teploty o 2-7 dní) alebo nedostatočné zvýšenie (o 0,2-0,3 °C) - pre horšiu funkciu žltého telieska, t.j. nedostatočná produkcia progesterónu. Falošne pozitívny výsledok (zvýšenie bazálnej teploty pri absencii žltého telieska) je možný pri akútnych a chronických infekciách, s niektorými zmenami v centrálnom nervovom systéme, sprevádzanými zvýšenou excitabilitou.
Symptóm "žiak" odráža množstvo a stav slizničnej sekrécie v cervikálnom kanáli, ktoré závisia od saturácie tela estrogénmi. Fenomén „zornice“ je založený na rozšírení vonkajšieho os cervikálneho kanála v dôsledku nahromadenia priehľadného sklovcového hlienu v ňom a hodnotí sa pri vyšetrovaní krčka maternice pomocou vaginálnych zrkadiel. V závislosti od závažnosti symptómu "žiaka" sa hodnotí v troch stupňoch: +, ++, +++.
Syntéza cervikálneho hlienu počas 1. fázy menštruačného cyklu sa zvyšuje a dosahuje maximum bezprostredne pred ovuláciou, čo je spojené s progresívnym zvýšením hladín estrogénu počas tohto obdobia. V predovulačných dňoch sa rozšírený vonkajší otvor cervikálneho kanála podobá zrenici (+++). V 2. fáze menštruačného cyklu sa množstvo estrogénu znižuje, progesterón sa tvorí prevažne vo vaječníkoch, takže množstvo hlienu klesá (+), pred menštruáciou úplne chýba (-). Test nemožno použiť na patologické zmeny na krčku maternice.
Symptóm kryštalizácie hlienu krčka maternice(fenomén "paprade") Pri vysychaní je najvýraznejší počas ovulácie, potom kryštalizácia postupne klesá a úplne chýba pred menštruáciou. Bodovo (od 1 do 3) sa hodnotí aj kryštalizácia na vzduchu vysušeného hlienu.
Symptóm napätia cervikálneho hlienu je priamo úmerná hladine estrogénu v ženskom tele. Na vykonanie testu sa hlien odstráni z cervikálneho kanála pomocou klieští, čeľuste nástroja sa pomaly pohybujú od seba, čím sa určuje stupeň napätia (vzdialenosť, pri ktorej sa hlien „láme“). K maximálnemu natiahnutiu hlienu krčka maternice (až 10-12 cm) dochádza v období najvyššej koncentrácie estrogénov – uprostred menštruačného cyklu, čo zodpovedá ovulácii.
Hlien môže byť negatívne ovplyvnený zápalovými procesmi v pohlavných orgánoch, ako aj hormonálnou nerovnováhou.
Karyopyknotický index(KPI). Vplyvom estrogénov dochádza k proliferácii buniek bazálnej vrstvy vrstevnatého dlaždicového epitelu vagíny, a preto sa v povrchovej vrstve zvyšuje počet keratinizujúcich (exfoliačných, odumierajúcich) buniek. Prvým štádiom bunkovej smrti sú zmeny v ich jadre (karyopyknóza). CPI je pomer počtu buniek s pyknotickým jadrom (t. j. keratinizujúcich) k celkovému počtu epiteliálnych buniek v nátere, vyjadrený v percentách. Na začiatku folikulárnej fázy menštruačného cyklu je CPI 20-40%, v predovulačných dňoch stúpa na 80-88%, čo súvisí s progresívnym zvyšovaním hladiny estrogénov. V luteálnej fáze cyklu hladina estrogénu klesá, preto CPI klesá na 20-25%. Kvantitatívne pomery bunkových elementov v náteroch vaginálnej sliznice teda umožňujú posúdiť saturáciu tela estrogénmi.
V súčasnosti sa najmä v programe mimotelového oplodnenia (IVF) pomocou dynamického ultrazvuku zisťuje dozrievanie folikulov, ovulácia a tvorba žltého telieska.
testovacie otázky
1. Popíšte normálny menštruačný cyklus.
2. Špecifikujte úrovne regulácie menštruačného cyklu.
3. Vymenujte princípy priamej a spätnej väzby.
4. Aké zmeny nastávajú vo vaječníkoch počas normálneho menštruačného cyklu?
5. Aké zmeny nastávajú v maternici počas normálneho menštruačného cyklu?
6. Vymenujte testy funkčnej diagnostiky.
Gynekológia: učebnica / B. I. Baisová a iné; vyd. G. M. Savelyeva, V. G. Breusenko. - 4. vydanie, prepracované. a dodatočné - 2011. - 432 s. : chorý.
Zoznam skratiek:
ADH - antidiuretický hormón
ACTH - kortikoliberín
aRG-GN - agonista hormónu uvoľňujúceho gonadotropín
LH - luteinizačný hormón
OP - oxyprogesterón
RG-GN - hormón uvoľňujúci gonadotropín
STH - somatoliberín
VEGF - vaskulárny endotelový rastový faktor
TSH - tyreotropný hormón (tyroliberín)
FSH - folikuly stimulujúci hormón
FGF - fibroplastický rastový faktor
Normálny menštruačný cyklus
Menštruácia- je to krvavý výtok z pohlavného traktu ženy, ktorý sa pravidelne vyskytuje v dôsledku odmietnutia funkčnej vrstvy endometria na konci dvojfázového menštruačného cyklu.
Komplex cyklických procesov, ktoré sa vyskytujú v ženskom tele a navonok sa prejavujú menštruáciou, sa nazýva menštruačný cyklus. Menštruácia začína ako odpoveď na zmenu hladiny steroidov produkovaných vaječníkmi.
Klinické príznaky normálneho menštruačného cykluTrvanie menštruačného cyklu v aktívnom reprodukčnom období ženy je v priemere 28 dní. Za normálnu sa považuje dĺžka cyklu 21 až 35 dní. Veľké intervaly sa pozorujú počas puberty a menopauzy, čo môže byť prejav anovulácie, ktorá sa v tomto období môže vyskytnúť najčastejšie.
Zvyčajne menštruácia trvá od 3 do 7 dní, množstvo stratenej krvi je zanedbateľné. Skrátenie alebo predĺženie menštruačného krvácania, ako aj výskyt slabej alebo silnej menštruácie môže slúžiť ako prejav celého radu gynekologických ochorení.
Charakteristiky normálneho menštruačného cyklu:
Trvanie: 28±7 dní;
Trvanie menštruačného krvácania: 4±2 dni;
Objem straty krvi počas menštruácie: 20-60 ml * ;
Priemerná strata železa: 16 mg
*
95 percent zdravých žien stratí pri každej menštruácii menej ako 60 ml krvi. Strata krvi viac ako 60-80 ml je kombinovaná s poklesom hemoglobínu, hematokritu a sérového železa.
Fyziológia menštruačného krvácania:
Bezprostredne pred menštruáciou sa vyvíja výrazný kŕč špirálovitých arteriol. Po rozšírení špirálových arteriol začína menštruačné krvácanie. Najprv je potlačená adhézia krvných doštičiek v endometriálnych cievach, ale potom, ako postupuje transudácia krvi, sú poškodené konce ciev uzavreté intravaskulárnymi trombami, ktoré pozostávajú z krvných doštičiek a fibrínu. 20 hodín po nástupe menštruácie, keď už bola väčšina endometria odtrhnutá, vzniká výrazný kŕč špirálovitých arteriol, vďaka čomu sa dosiahne hemostáza. Regenerácia endometria začína 36 hodín po začiatku menštruácie, napriek tomu, že odmietnutie endometria ešte nie je úplne dokončené.
Regulácia menštruačného cyklu je komplexný neurohumorálny mechanizmus, ktorý sa uskutočňuje za účasti 5 hlavných článkov regulácie. Patria sem: mozgová kôra, subkortikálne centrá (hypotalamus), hypofýza, pohlavné žľazy, periférne orgány a tkanivá (maternica, vajíčkovody, vagína, mliečne žľazy, vlasové folikuly, kosti, tukové tkanivo). Posledne menované sa nazývajú cieľové orgány, kvôli prítomnosti receptorov, ktoré sú citlivé na pôsobenie hormónov, ktoré vaječník produkuje počas menštruačného cyklu. Cytosolové receptory - receptory cytoplazmy, majú prísnu špecifickosť pre estradiol, progesterón, testosterón, zatiaľ čo jadrové receptory môžu byť akceptormi molekúl ako inzulín, glukagón, aminopeptidy.
Receptory pre pohlavné hormóny sa nachádzajú vo všetkých štruktúrach reprodukčného systému, ako aj v centrálnom nervovom systéme, koži, tukovom a kostnom tkanive a mliečnej žľaze. Voľná molekula steroidného hormónu je zachytená špecifickým cytosolovým receptorom proteínovej povahy, výsledný komplex je translokovaný do bunkového jadra. V jadre sa objavuje nový komplex s jadrovým proteínovým receptorom; tento komplex sa viaže na chromatín, ktorý reguluje transkripciu mRNA a podieľa sa na syntéze špecifického tkanivového proteínu. Intracelulárny mediátor - cyklická kyselina adenozínmonofosforečná (cAMP) reguluje metabolizmus v bunkách cieľového tkaniva v súlade s potrebami organizmu v reakcii na účinky hormónov. Prevažná časť steroidných hormónov (asi 80 % je v krvi a transportuje sa vo viazanej forme. Ich transport zabezpečujú špeciálne proteíny - steroidy viažuce globulíny a nešpecifické transportné systémy (albumíny a erytrocyty). Vo viazanej forme steroidy sú neaktívne, preto globulíny, albumíny a erytrocyty možno považovať za akýsi nárazníkový systém, ktorý riadi prístup steroidov k receptorom cieľových buniek.
Cyklické funkčné zmeny vyskytujúce sa v ženskom tele možno podmienečne rozdeliť na zmeny v systéme hypotalamus-hypofýza-vaječníky (ovariálny cyklus) a v maternici, predovšetkým v jej sliznici (cyklus maternice).
Spolu s tým sa spravidla vyskytujú cyklické posuny vo všetkých orgánoch a systémoch ženy, najmä v centrálnom nervovom systéme, kardiovaskulárnom systéme, termoregulačnom systéme, metabolických procesoch atď.
Hypotalamus
Hypotalamus je časť mozgu umiestnená nad optickou chiazmou a tvoriaca spodok tretej komory. Ide o starú a stabilnú zložku centrálneho nervového systému, ktorej všeobecná organizácia sa počas evolúcie človeka len málo zmenila. Štrukturálne a funkčne súvisí hypotalamus s hypofýzou. Existujú tri oblasti hypotalamu: predná, zadná a stredná. Každá oblasť je tvorená jadrami - akumuláciami tiel neurónov určitého typu.
Okrem hypofýzy ovplyvňuje hypotalamus limbický systém (amygdala, hipokampus), talamus a mostík. Tieto oddelenia tiež priamo alebo nepriamo ovplyvňujú hypotalamus.
Hypotalamus vylučuje liberíny a statíny. Tento proces je regulovaný hormónmi, ktoré uzatvárajú tri slučky spätnej väzby: dlhú, krátku a ultrakrátku. Dlhú spätnú väzbu poskytujú cirkulujúce pohlavné hormóny, ktoré sa viažu na zodpovedajúce receptory v hypotalame, krátku: hormóny adenohypofýzy, ultrakrátku: liberíny a statíny. Liberíny a statíny regulujú činnosť adenohypofýzy. Gonadoliberín stimuluje sekréciu LH a FSH, kortikoliberínu - ACTH, somatoliberínu (STG), tyreoliberínu (TSH). Okrem liberínov a statínov sa v hypotalame syntetizuje antidiuretický hormón a oxytocín. Tieto hormóny sú transportované do neurohypofýzy, odkiaľ sa dostávajú do krvného obehu.
Na rozdiel od kapilár iných oblastí mozgu sú kapiláry lievika hypotalamu fenestrované. Tvoria primárnu kapilárnu sieť portálového systému.
V 70-80 rokoch. bola vykonaná séria experimentálnych štúdií na opiciach, ktoré umožnili identifikovať rozdiely vo funkcii neurosekrečných štruktúr hypotalamu primátov a hlodavcov. U primátov a ľudí sú oblúkovité jadrá mediobazálneho hypotalamu jediným miestom pre tvorbu a uvoľňovanie RG-LH, ktorý je zodpovedný za gonadotropnú funkciu hypofýzy. Sekrécia RG-LH je geneticky naprogramovaná a prebieha v určitom pulzujúcom rytme s frekvenciou približne raz za hodinu. Tento rytmus sa nazýva cirhorálny (hodina-tá). Oblasť oblúkových jadier hypotalamu sa nazýva oblúkový oscilátor. Cirkorálny charakter sekrécie RG-LH bol potvrdený jeho priamym stanovením v krvi portálneho systému hypofýzovej stopky a jugulárnej žily u opíc a v krvi žien s ovulačným cyklom.
Hormóny hypotalamu
Uvoľňujúci hormón LH bol izolovaný, syntetizovaný a podrobne opísaný. Doteraz nebolo možné izolovať a syntetizovať foliberín. RG-LH a jeho syntetické analógy majú schopnosť stimulovať uvoľňovanie LH a FSH z prednej hypofýzy, preto sa v súčasnosti akceptuje jeden termín pre hypotalamické gonadotropné liberíny - gonadotropín uvoľňujúci hormón (RG-GN).
Gonadoliberín stimuluje sekréciu FSH a LH. Je to dekapeptid vylučovaný neurónmi nucleus infundibulum. Gonadoliberín sa nevylučuje neustále, ale v pulznom režime. Je veľmi rýchlo zničený proteázami (polčas rozpadu je 2–4 min), takže jeho impulzy musia byť pravidelné. Frekvencia a amplitúda emisií GnRH sa mení počas menštruačného cyklu. Folikulárna fáza je charakterizovaná častými výkyvmi v malej amplitúde hladiny gonadoliberínu v krvnom sére. Ku koncu folikulárnej fázy sa frekvencia a amplitúda oscilácií zvyšuje a potom klesá počas luteálnej fázy.
Hypofýza
V hypofýze sú dva laloky: predná - adenohypofýza a zadná - neurohypofýza. Neurohypofýza je neurogénneho pôvodu a predstavuje pokračovanie lievika hypotalamu. Neurohypofýza je zásobovaná krvou z dolných hypofýzových tepien. Adenohypofýza sa vyvíja z ektodermy Rathkeho vačku, preto pozostáva zo žľazového epitelu a nemá priame spojenie s hypotalamom. Liberíny a statíny syntetizované v hypotalame vstupujú do adenohypofýzy cez špeciálny portálový systém. Je hlavným zdrojom krvného zásobenia adenohypofýzy. Krv vstupuje do portálneho systému hlavne cez horné hypofýzové tepny. V oblasti lievika hypotalamu tvoria primárnu kapilárnu sieť portálneho systému, z ktorej sa vytvárajú portálne žily, ktoré vstupujú do adenohypofýzy a dávajú vznik sekundárnej kapilárnej sieti. Je možný spätný tok krvi cez portálový systém. Vlastnosti krvného zásobenia a neprítomnosť hematoencefalickej bariéry v lieviku hypotalamu poskytujú obojsmerné spojenie medzi hypotalamom a hypofýzou. V závislosti od farbenia hematoxylínom a eozínom sa sekrečné bunky adenohypofýzy delia na chromofilné (acidofilné) a bazofilné (chromofóbne). Acidofilné bunky vylučujú rastový hormón a prolaktín, bazofilné bunky - FSH, LH, TSH, ACTH
hormóny hypofýzy
Adenohypofýza produkuje GH, prolaktín, FSH, LH, TSH a ACTH. FSH a LH regulujú sekréciu pohlavných hormónov, TSH - sekréciu hormónov štítnej žľazy, ACTH - sekréciu hormónov kôry nadobličiek. STH stimuluje rast, pôsobí anabolicky. Prolaktín stimuluje rast mliečnych žliaz počas tehotenstva a laktácie po pôrode.
LH a FSH sú syntetizované gonadotropnými bunkami adenohypofýzy a hrajú dôležitú úlohu pri vývoji ovariálnych folikulov. Štruktúrne sú klasifikované ako glykoproteíny. FSH stimuluje rast folikulov, proliferáciu granulóznych buniek, vyvoláva tvorbu LH receptorov na povrchu granulózových buniek. Vplyvom FSH sa zvyšuje obsah aromatázy v dozrievajúcom folikule. LH stimuluje tvorbu androgénov (estrogénových prekurzorov) v bunkách theca, spolu s FSH podporuje ovuláciu a stimuluje syntézu progesterónu v luteinizovaných granulózových bunkách ovulovaného folikulu.
Sekrécia LH a FSH je variabilná a modulovaná ovariálnymi hormónmi, najmä estrogénom a progesterónom.
Nízka hladina estrogénu má teda supresívny účinok na LH, zatiaľ čo vysoká hladina stimuluje jeho produkciu hypofýzou. V neskorej folikulárnej fáze sú hladiny estrogénu v sére dosť vysoké, efekt pozitívnej spätnej väzby sa strojnásobí, čo prispieva k vytvoreniu predovulačného vrcholu LH. A naopak, pri terapii kombinovanou antikoncepciou je hladina estrogénu v krvnom sére v medziach, ktoré určujú negatívnu spätnú väzbu, čo vedie k zníženiu obsahu gonadotropínov.
Mechanizmus pozitívnej spätnej väzby vedie k zvýšeniu koncentrácie a produkcie RG-GN v receptoroch.
Na rozdiel od účinku estrogénov majú nízke hladiny progesterónu pozitívnu spätnú väzbu na sekréciu LH a FSH hypofýzou. Tieto stavy existujú tesne pred ovuláciou a vedú k uvoľneniu FSH. Vysoká hladina progesterónu, ktorá je zaznamenaná v luteálnej fáze, znižuje produkciu gonadotropínov hypofýzou. Malé množstvo progesterónu stimuluje uvoľňovanie gonadotropínov na úrovni hypofýzy. Negatívny spätnoväzbový efekt progesterónu sa prejavuje znížením produkcie RG-GN a znížením citlivosti na RG-GN na úrovni hypofýzy. Pozitívna spätná väzba progesterónu sa vyskytuje na hypofýze a zahŕňa zvýšenú citlivosť na RH-GN. Estrogény a progesterón nie sú jedinými hormónmi, ktoré ovplyvňujú sekréciu gonadotropínov hypofýzou. Rovnaký účinok majú hormóny inhibín a aktivín. Inhibín potláča sekréciu FSH hypofýzy, zatiaľ čo aktivín ju stimuluje.
Prolaktín je polypeptid pozostávajúci zo 198 aminokyselinových zvyškov, syntetizovaný laktotropnými bunkami adenohypofýzy. Sekrécia prolaktínu je riadená dopamínom. Je syntetizovaný v hypotalame a inhibuje sekréciu prolaktínu. Prolaktín má rôzne účinky na telo ženy. Jeho hlavnou biologickou úlohou je rast mliečnych žliaz a regulácia laktácie. Má tiež tuk mobilizujúci účinok a má hypotenzný účinok. Zvýšenie sekrécie prolaktínu je jednou z bežných príčin neplodnosti, pretože zvýšenie jeho hladiny v krvi inhibuje steroidogenézu vo vaječníkoch a vývoj folikulov.
Oxytocín- peptid pozostávajúci z 9 aminokyselinových zvyškov. Tvorí sa v neurónoch veľkej bunkovej časti paraventrikulárnych jadier hypotalamu. Hlavnými cieľmi oxytocínu u ľudí sú hladké svalové vlákna maternice a myoepiteliálne bunky mliečnych žliaz.
Antidiuretický hormón(ADH) je peptid pozostávajúci z 9 aminokyselinových zvyškov. Syntetizovaný v neurónoch supraoptického jadra hypotalamu. Hlavnou funkciou ADH je regulácia BCC, krvného tlaku a osmolality plazmy.
Ovariálny cyklus
Vaječníky prechádzajú tromi fázami menštruačného cyklu:
- folikulárna fáza;
- ovulácia;
- luteálna fáza.
Folikulárna fáza:
Jedným z vrcholov folikulárnej fázy menštruačného cyklu je vývoj vajíčka. Vaječník ženy je komplexný orgán pozostávajúci z mnohých zložiek, v dôsledku ktorých sa v dôsledku interakcie vylučujú pohlavné steroidné hormóny a v reakcii na cyklickú sekréciu gonadotropínov sa vytvára vajíčko pripravené na oplodnenie.
Steroidogenéza
Hormonálna aktivita od preantrálneho po periovulačný folikul bola opísaná ako teória „dve bunky, dva gonadotropíny“. Steroidogenéza sa vyskytuje v dvoch bunkách folikulu: theca a granulózových bunkách. V bunkách theca stimuluje LH produkciu androgénov z cholesterolu. V granulózových bunkách FSH stimuluje premenu výsledných androgénov na estrogény (aromatizácia). Okrem aromatizačného účinku je FSH zodpovedný aj za proliferáciu granulóznych buniek. Hoci sú známe ďalšie mediátory vo vývoji ovariálnych folikulov, táto teória je hlavnou teóriou pre pochopenie procesov vyskytujúcich sa vo ovariálnych folikuloch. Ukázalo sa, že oba hormóny sú potrebné pre normálny cyklus s dostatočnou hladinou estrogénu.
Produkcia androgénov vo folikuloch môže tiež regulovať vývoj preantrálneho folikulu. Nízka hladina androgénov podporuje proces aromatizácie, teda zvyšuje produkciu estrogénu a naopak vysoká hladina brzdí proces aromatizácie a spôsobuje atréziu folikulu. Rovnováha FSH a LH je nevyhnutná pre skorý vývoj folikulov. Optimálnou podmienkou pre počiatočné štádium vývoja folikulov je nízka hladina LH a vysoký FSH, ku ktorému dochádza na začiatku menštruačného cyklu. Ak je hladina LH vysoká, bunky theca produkujú veľké množstvo androgénov, čo spôsobuje folikulárnu atréziu.
Výber dominantného folikulu
Rast folikulu je sprevádzaný sekréciou pohlavných steroidných hormónov pod vplyvom LH a FSH. Tieto gonadotropíny chránia skupinu preantrálnych folikulov pred atréziou. Normálne sa však len jeden z týchto folikulov vyvinie do predovulačného folikulu, ktorý sa potom uvoľní a stane sa dominantným.
Dominantný folikul v strednej folikulárnej fáze je najväčší a najrozvinutejší vo vaječníku. Už v prvých dňoch menštruačného cyklu má priemer 2 mm a do 14 dní v čase ovulácie sa zväčší v priemere na 21 mm. Počas tejto doby sa objem folikulárnej tekutiny zväčší 100-násobne, počet granulóznych buniek lemujúcich bazálnu membránu sa zvýši z 0,5x106 na 50x106. Tento folikul má najvyššiu aromatizačnú aktivitu a najvyššiu koncentráciu FSH-indukovaných LH receptorov, takže dominantný folikul vylučuje najvyššie množstvo estradiolu a inhibínu. Ďalej inhibín zvyšuje syntézu androgénov pod vplyvom LH, ktorý je substrátom pre syntézu estradiolu.
Na rozdiel od hladiny FSH, ktorá klesá so zvyšujúcou sa koncentráciou estradiolu, hladina LH stále stúpa (pri nízkych koncentráciách estradiol inhibuje sekréciu LH). Práve dlhodobá estrogénová stimulácia pripravuje ovulačný vrchol LH. Súčasne sa dominantný folikul pripravuje na ovuláciu: pod lokálnym pôsobením estrogénov a FSH sa zvyšuje počet LH receptorov na granulóznych bunkách. Uvoľňovanie LH vedie k ovulácii, tvorbe žltého telieska a zvýšeniu sekrécie progesterónu. K ovulácii dochádza 10-12 hodín po vrchole LH alebo 32-35 hodín po začiatku vzostupu jeho hladiny. Zvyčajne ovuluje iba jeden folikul.
Počas selekcie folikulov hladiny FSH klesajú v reakcii na negatívne účinky estrogénu, takže dominantný folikul je jediný, ktorý sa naďalej vyvíja s klesajúcimi hladinami FSH.
Ovariálne-hypofyzárne spojenie je rozhodujúce pri výbere dominantného folikulu a pri vzniku atrézie zvyšných folikulov.
inhibín a aktivín
K rastu a vývoju vajíčka, fungovaniu žltého telieska dochádza prostredníctvom interakcie autokrinných a parakrinných mechanizmov. Treba poznamenať dva folikulárne hormóny, ktoré zohrávajú významnú úlohu v steroidogenéze - inhibín a aktivín.
Inhibín je peptidový hormón produkovaný granulóznymi bunkami rastúcich folikulov, ktorý znižuje produkciu FSH. Okrem toho ovplyvňuje syntézu androgénov vo vaječníku. Inhibín ovplyvňuje folikulogenézu nasledujúcim spôsobom: znížením FSH na úroveň, pri ktorej sa vyvíja iba dominantný folikul.
Aktivín je peptidový hormón produkovaný v granulóznych bunkách folikulov a hypofýzy. Podľa niektorých autorov je aktivín produkovaný aj placentou. Aktivín zvyšuje produkciu FSH hypofýzou, zvyšuje väzbu FSH na bunky granulózy.
Rastové faktory podobné inzulínu
Inzulínu podobné rastové faktory (IGF-1 a IGF-2) sa syntetizujú v pečeni pod vplyvom rastového hormónu a prípadne v granulóznych bunkách folikulov pôsobia ako parakrinné regulátory. Pred ovuláciou sa obsah IGF-1 a IGF-2 vo folikulárnej tekutine zvyšuje v dôsledku zvýšenia množstva samotnej tekutiny v dominantnom folikule. IGF-1 sa podieľa na syntéze estradiolu. IGF-2 (epidermálny) inhibuje syntézu steroidov vo vaječníkoch.
Ovulácia:
Ovulačný vrchol LH vedie k zvýšeniu koncentrácie prostaglandínov a proteázovej aktivity vo folikule. Samotný proces ovulácie je pretrhnutie bazálnej membrány dominantného folikulu a krvácanie zo zničených kapilár obklopujúcich bunky theca. Zmeny v stene predovulačného folikulu, ktoré zabezpečujú jeho stenčenie a prasknutie, sa vyskytujú pod vplyvom enzýmu kolagenázy; určitú úlohu zohrávajú aj prostaglandíny obsiahnuté vo folikulárnej tekutine, proteolytické enzýmy tvorené v granulózových bunkách, oxytopín a relaxín. V dôsledku toho sa v stene folikulu vytvorí malý otvor, cez ktorý sa vajíčko pomaly uvoľňuje. Priame merania ukázali, že tlak vo folikule sa počas ovulácie nezvyšuje.
Na konci folikulárnej fázy pôsobí FSH na LH receptory v granulóznych bunkách. Estrogény sú povinným kofaktorom tohto účinku. Ako sa dominantný folikul vyvíja, produkcia estrogénu sa zvyšuje. V dôsledku toho je produkcia estrogénov dostatočná na dosiahnutie sekrécie LH hypofýzou, čo vedie k zvýšeniu jeho hladiny. K zvýšeniu dochádza najskôr veľmi pomaly (od 8. do 12. dňa cyklu), potom rýchlo (po 12. dni cyklu). Počas tejto doby LH aktivuje luteinizáciu granulózových buniek v dominantnom folikule. Tak sa uvoľňuje progesterón. Progesterón ďalej zvyšuje účinok estrogénov na sekréciu hypofýzového LH, čo vedie k zvýšeniu jeho hladiny.
K ovulácii dochádza do 36 hodín od začiatku nárastu LH. Stanovenie nárastu LH je jednou z najlepších metód na určenie ovulácie a vykonáva sa pomocou zariadenia "detektor ovulácie".
Periovulačný vrchol FSH pravdepodobne nastáva v dôsledku pozitívneho účinku progesterónu. Okrem zvýšenia LH, FSH a estrogénov dochádza aj k zvýšeniu sérových hladín androgénov počas ovulácie. Tieto androgény sa uvoľňujú v dôsledku stimulačného účinku LH na bunky theca, najmä v nedominantnom folikule.
Nárast androgénov má vplyv na zvýšené libido, čo potvrdzuje, že toto obdobie je u žien najplodnejšie.
Hladiny LH stimulujú meiózu po vstupe spermií do vajíčka. Keď sa oocyt uvoľní z vaječníka počas ovulácie, stena folikulu je zničená. Toto je regulované LH, FSH a progesterónom, ktoré stimulujú aktivitu proteolytických enzýmov, ako sú aktivátory plazminogénu (uvoľňujúce plazmín, ktorý stimuluje aktivitu kolagenázy) a prostaglandíny. Prostaglandíny nielen zvyšujú aktivitu proteolytických enzýmov, ale prispievajú aj k vzniku zápalovej reakcie v stene folikulu a stimulujú činnosť hladkého svalstva, čo prispieva k uvoľneniu oocytu.
Význam prostaglandínov v procese ovulácie bol dokázaný štúdiami, ktoré naznačujú, že zníženie uvoľňovania prostaglandínov môže viesť k oneskoreniu uvoľňovania oocytu z vaječníka počas normálnej steroidogenézy (syndróm nevyvíjajúceho sa luteinizovaného folikulu - SNLF). Keďže SNLF je často príčinou neplodnosti, ženám, ktoré chcú otehotnieť, sa odporúča vyhýbať sa užívaniu syntetizovaných inhibítorov prostaglandínov.
luteálna fáza:
Štruktúra corpus luteum
Po uvoľnení vajíčka z vaječníka tvoriace sa kapiláry rýchlo rastú do dutiny folikulu; granulózne bunky podliehajú luteinizácii: zvýšeniu cytoplazmy v nich a tvorbe lipidových inklúzií. Granulózne bunky a tekocyty tvoria žlté teliesko, hlavný regulátor luteálnej fázy menštruačného cyklu. Bunky, ktoré vytvorili stenu folikulu, akumulujú lipidy a žltý pigment luteín a začnú vylučovať progesterón, estradiol-2 a inhibín. Výkonná vaskulárna sieť prispieva k vstupu hormónov žltého telieska do systémového obehu. Plnohodnotné žlté teliesko sa vyvinie až vtedy, keď sa v predovulačnom folikule vytvorí primeraný počet granulóznych buniek s vysokým obsahom LH receptorov. K zvýšeniu veľkosti žltého telieska po ovulácii dochádza najmä v dôsledku zväčšenia veľkosti buniek granulózy, zatiaľ čo ich počet sa nezvyšuje v dôsledku absencie mitóz. U ľudí corpus luteum vylučuje nielen progesterón, ale aj estradiol a androgény. Mechanizmy regresie corpus luteum nie sú dobre známe. Je známe, že prostaglandíny majú luteolytický účinok.

Ryža. Ultrazvukový obraz "rozkvitnutého" žltého telieska počas tehotenstva 6 týždňov. 4 dni. Režim energetického mapovania.
Hormonálna regulácia luteálnej fázy
Ak nedôjde k otehotneniu, dôjde k involúcii žltého telieska. Tento proces je regulovaný mechanizmom negatívnej spätnej väzby: hormóny (progesterón a estradiol) vylučované žltým telom pôsobia na gonadotropné bunky hypofýzy, čím potláčajú sekréciu FSH a LH. Inhibín tiež inhibuje sekréciu FSH. Pokles hladín FSH, ako aj lokálne pôsobenie progesterónu bráni rozvoju skupiny primordiálnych folikulov.
Existencia corpus luteum závisí od úrovne sekrécie LH. Keď sa zníži, zvyčajne 12-16 dní po ovulácii, dochádza k involúcii žltého telieska. Na jeho mieste sa vytvorí biele telo. Mechanizmus involúcie nie je známy. S najväčšou pravdepodobnosťou je to spôsobené parakrinnými vplyvmi. Pri involúcii žltého telieska hladiny estrogénu a progesterónu klesajú, čo vedie k zvýšenej sekrécii gonadotropných hormónov. Keď sa obsah FSH a LH zvyšuje, začína sa vyvíjať nová skupina folikulov.
Ak došlo k oplodneniu, existenciu žltého telieska a sekréciu progesterónu podporuje choriový gonadotropín. Implantácia embrya teda vedie k hormonálnym zmenám, ktoré zachovávajú žlté teliesko.
Trvanie luteálnej fázy u väčšiny žien je konštantné a je približne 14 dní.
Ovariálne hormóny
Komplexný proces biosyntézy steroidov končí tvorbou estradiolu, testosterónu a progesterónu. Steroidy produkujúce tkanivá vaječníkov sú granulózne bunky vystielajúce dutinu folikulu, bunky vnútornej théky a v oveľa menšej miere stróma. Bunky granulózy a bunky theca sa synergicky podieľajú na syntéze estrogénov, bunky tekálnej membrány sú hlavným zdrojom androgénov, ktoré sa v malom množstve tvoria aj v stróme; progesterón sa syntetizuje v bunkách theca a granulózových bunkách.
Vo vaječníku sa 60-100 mcg estradiolu (E2) vylučuje v skorej folikulárnej fáze menštruačného cyklu, 270 mcg v luteálnej fáze a 400-900 mcg denne v čase ovulácie. Asi 10% E2 je aromatizovaných vo vaječníkoch z testosterónu. Množstvo estrónu vytvoreného vo včasnej folikulárnej fáze je 60-100 mcg, v čase ovulácie sa jeho syntéza zvýši na 600 mcg za deň. Iba polovica množstva estrónu sa produkuje vo vaječníku. Druhá polovica je aromatizovaná na E2. Estriol je neaktívny metabolit estradiolu a estrónu.
Progesterón sa produkuje vo vaječníkoch v dávke 2 mg/deň počas folikulárnej fázy a 25 mg/deň počas luteálnej fázy menštruačného cyklu. V procese metabolizmu sa progesterón vo vaječníku mení na 20-dehydroprogesterón, ktorý má relatívne nízku biologickú aktivitu.
Vo vaječníku sa syntetizujú tieto androgény: androstendión (prekurzor testosterónu) v množstve 1,5 mg/deň (rovnaké množstvo androstendiónu sa tvorí v nadobličkách). Z androstendiónu sa tvorí asi 0,15 mg testosterónu, približne rovnaké množstvo sa tvorí v nadobličkách.
Stručný prehľad procesov prebiehajúcich vo vaječníkoch
Folikulárna fáza:
LH stimuluje produkciu androgénov v bunkách theca.
FSH stimuluje produkciu estrogénu v granulózových bunkách.
Dominantným sa stáva najviac rozvinutý folikul v strede folikulárnej fázy.
Zvyšujúca sa produkcia estrogénov a inhibínu v dominantnom folikule potláča uvoľňovanie FSH hypofýzou.
Pokles hladín FSH spôsobuje atréziu všetkých folikulov okrem dominantného.
Ovulácia:
FSH indukuje LH receptory.
Proteolytické enzýmy vo folikule vedú k deštrukcii jeho steny a uvoľneniu oocytu.
luteálna fáza:
Žlté teliesko sa tvorí z buniek granulózy a théky zachovaných po ovulácii.
Dominantným hormónom je progesterón, vylučovaný žltým telom. Pri absencii tehotenstva nastáva luteolýza 14 dní po ovulácii.
cyklus maternice
Endometrium pozostáva z dvoch vrstiev: funkčnej a bazálnej. Funkčná vrstva mení svoju štruktúru pôsobením pohlavných hormónov a ak nedôjde k otehotneniu, je počas menštruácie odmietnutá.
Proliferatívna fáza:
Za začiatok menštruačného cyklu sa považuje 1. deň menštruácie. Na konci menštruácie je hrúbka endometria 1-2 mm. Endometrium pozostáva takmer výlučne z bazálnej vrstvy. Žľazy sú úzke, rovné a krátke, lemované nízkym cylindrickým epitelom, cytoplazma stromálnych buniek je takmer rovnaká. Keď sa hladina estradiolu zvýši, vytvorí sa funkčná vrstva: endometrium sa pripravuje na implantáciu embrya. Žľazy sa predlžujú a stávajú sa kľukatými. Zvyšuje sa počet mitóz. S proliferáciou sa výška epitelových buniek zvyšuje a samotný epitel z jedného radu sa v čase ovulácie stáva viacradovým. Stróma je edematózna a uvoľnená, zväčšujú sa v nej jadrá buniek a objem cytoplazmy. Cievy sú stredne kľukaté.
sekrečná fáza:
Normálne nastáva ovulácia na 14. deň menštruačného cyklu. Sekrečná fáza je charakterizovaná vysokými hladinami estrogénu a progesterónu. Po ovulácii sa však počet estrogénových receptorov v bunkách endometria znižuje. Postupne sa inhibuje proliferácia endometria, znižuje sa syntéza DNA a znižuje sa počet mitóz. Progesterón má teda prevládajúci účinok na endometrium v sekrečnej fáze.
V žľazách endometria sa objavujú vakuoly obsahujúce glykogén, ktoré sa detegujú pomocou PAS reakcie. Na 16. deň cyklu sú tieto vakuoly pomerne veľké, nachádzajú sa vo všetkých bunkách a nachádzajú sa pod jadrami. Na 17. deň sa jadrá, vytlačené vakuolami, nachádzajú v centrálnej časti bunky. Na 18. deň sú vakuoly v apikálnej časti a jadrá v bazálnej časti buniek, glykogén sa začína uvoľňovať do lumen žliaz apokrinnou sekréciou. Najlepšie podmienky na implantáciu sú vytvorené na 6.-7.deň po ovulácii, t.j. na 20-21 deň cyklu, kedy je sekrečná aktivita žliaz maximálna.
V 21. deň cyklu začína deciduálna reakcia endometriálnej strómy. Špirálové tepny sú ostro kľukaté, neskôr v dôsledku zníženia edému strómy sú zreteľne viditeľné. Najprv sa objavia deciduálne bunky, ktoré postupne vytvárajú zhluky. Na 24. deň cyklu tieto nahromadenia tvoria perivaskulárne eozinofilné mufy. Na 25. deň sa vytvárajú ostrovčeky deciduálnych buniek. Do 26. dňa cyklu sa deciduálna reakcia stáva počtom neutrofilov, ktoré tam migrujú z krvi. Neutrofilná infiltrácia je nahradená nekrózou funkčnej vrstvy endometria.
menštruácia:
Ak nedôjde k implantácii, žľazy prestanú produkovať tajomstvo a začnú sa degeneratívne zmeny vo funkčnej vrstve endometria. Bezprostredným dôvodom jeho odmietnutia je prudký pokles obsahu estradiolu a progesterónu v dôsledku involúcie žltého telieska. V endometriu sa znižuje venózny odtok a dochádza k vazodilatácii. Potom dochádza k zúženiu tepien, čo vedie k ischémii a poškodeniu tkaniva a funkčnej strate endometria. Potom dochádza ku krvácaniu z fragmentov arteriol zostávajúcich v bazálnej vrstve endometria. Menštruácia sa zastaví zúžením tepien, obnoví sa endometrium. Zastavenie krvácania v cievach endometria sa teda líši od hemostázy v iných častiach tela.
Krvácanie sa spravidla zastaví v dôsledku hromadenia krvných doštičiek a ukladania fibrínu, čo vedie k zjazveniu. V endometriu môže zjazvenie viesť k strate jeho funkčnej aktivity (Ashermanov syndróm). Aby sa predišlo týmto následkom, je potrebný alternatívny systém hemostázy. Cievna kontrakcia je mechanizmus na zastavenie krvácania v endometriu. Zjazvenie sa zároveň minimalizuje fibrinolýzou, ktorá ničí krvné zrazeniny. Neskôr obnovenie endometria a tvorba nových krvných ciev (angiogenéza) vedie k ukončeniu krvácania do 5-7 dní od začiatku menštruačného cyklu.
Účinok vysadenia estrogénu a progesterónu na menštruáciu je dobre definovaný, ale úloha parakrinných mediátorov zostáva nejasná. Vazokonstriktory: prostaglandín F2a, endotel-1 a faktor aktivujúci doštičky (TAF) sa môžu vytvárať v endometriu a podieľať sa na vazokonstrikcii. Prispievajú tiež k nástupu menštruácie a ďalšej kontrole nad ňou. Tieto mediátory môžu byť regulované pôsobením vazodilatátorov, ako je prostaglandín E2, prostacyklín, oxid dusnatý, ktoré sú produkované endometriom. Prostaglandín F2a má výrazný vazokonstrikčný účinok, zvyšuje arteriálny spazmus a ischémiu endometria, spôsobuje kontrakcie myometria, čo na jednej strane znižuje prietok krvi a na druhej strane pomáha odstraňovať odmietnuté endometrium.
Oprava endometria zahŕňa glandulárnu a stromálnu regeneráciu a angiogenézu. Vaskulárny endoteliálny rastový faktor (VEGF) a fibroplastický rastový faktor (FGF) sa nachádzajú v endometriu a sú silnými činiteľmi angiogenézy. Zistilo sa, že estrogénom produkovaná žľazová a stromálna regenerácia je zvýšená pod vplyvom epidermálnych rastových faktorov (EGF). Rastové faktory, ako je transformujúci rastový faktor (TGF) a interleukíny, najmä interleukín-1 (IL-1), sú veľmi dôležité.
Stručný prehľad procesov prebiehajúcich v endometriu
menštruácia:
Hlavnú úlohu na začiatku menštruácie zohráva kŕč arteriol.
Funkčná vrstva endometria (horná, tvoriaca 75 % hrúbky) je odmietnutá.
Menštruácia sa zastaví v dôsledku vazospazmu a obnovy endometria. Fibrinolýza zabraňuje tvorbe zrastov.
Proliferatívna fáza:
Je charakterizovaná estrogénom indukovanou proliferáciou žliaz a strómy.
sekrečná fáza:
Je charakterizovaná sekréciou žliaz vyvolanou progesterónom.
V neskorej sekrečnej fáze je navodená decidualizácia.
Decidualizácia je nezvratný proces. Pri absencii tehotenstva dochádza v endometriu k apoptóze, po ktorej nasleduje menštruácia.
Reprodukčný systém je teda supersystém, ktorého funkčný stav je určený reverznou aferentáciou jeho základných subsystémov. Prideliť: dlhú spätnú väzbu medzi hormónmi vaječníka a jadrami hypotalamu; medzi ovariálnymi hormónmi a hypofýzou; krátka slučka medzi prednou hypofýzou a hypotalamom; ultrashort medzi RG-LH a neurocytmi (nervovými bunkami) hypotalamu.
Spätná väzba od sexuálne zrelej ženy je negatívna aj pozitívna. Príkladom negatívnej asociácie je zvýšenie uvoľňovania LH z prednej hypofýzy v reakcii na nízke hladiny estradiolu v skorej folikulárnej fáze cyklu. Príkladom pozitívnej spätnej väzby je uvoľňovanie LH a FSH v reakcii na ovulačné maximum estradiolu v krvi. Podľa mechanizmu negatívnej spätnej väzby sa tvorba RG-LH zvyšuje s poklesom hladiny LH v bunkách prednej hypofýzy.
Zhrnutie
GnRH je syntetizovaný neurónmi infundibulum nucleus, potom vstupuje do portálneho systému hypofýzy a cez ňu vstupuje do adenohypofýzy. Sekrécia GnRH prebieha impulzívne.
Skoré štádium vývoja skupiny primordiálnych folikulov je nezávislé od FSH.
Ako sa žlté teliesko involuje, sekrécia progesterónu a inhibínu klesá a hladiny FSH stúpajú.
FSH stimuluje rast a vývoj skupiny primordiálnych folikulov a ich sekréciu estrogénov.
Estrogény pripravujú maternicu na implantáciu stimuláciou proliferácie a diferenciácie funkčnej vrstvy endometria a spolu s FSH podporujú vývoj folikulov.
Podľa dvojbunkovej teórie syntézy pohlavných hormónov LH stimuluje syntézu androgénov v thekocytoch, ktoré sa potom v granulóznych bunkách vplyvom FSH premieňajú na estrogény.
Zvýšenie koncentrácie estradiolu mechanizmom negatívnej spätnej väzby, slučkou
ktorý sa uzatvára v hypofýze a hypotalame, potláča sekréciu FSH.
Folikul, ktorý bude ovulovať v danom menštruačnom cykle, sa nazýva dominantný folikul. Na rozdiel od iných folikulov, ktoré začali rásť, nesie väčší počet FSH receptorov a syntetizuje viac estrogénu. To mu umožňuje rozvíjať sa napriek poklesu hladín FSH.
Dostatočná estrogénová stimulácia poskytuje ovulačný vrchol LH. Tá zase spôsobuje ovuláciu, tvorbu žltého telieska a vylučovanie progesterónu.
Fungovanie corpus luteum závisí od hladiny LH. S jeho poklesom dochádza k involúcii žltého telieska. K tomu zvyčajne dochádza na 12-16 deň po ovulácii.
Ak došlo k oplodneniu, existenciu žltého telieska podporuje choriový gonadotropín. Žlté teliesko naďalej vylučuje progesterón, ktorý je nevyhnutný na udržanie tehotenstva v počiatočných štádiách.
Regulácia menštruačného cyklu prebieha na piatich rôznych úrovniach.: od mozgovej kôry po hlavný orgán - maternicu.
Aby ste pochopili, ako to chodí predstavte si hodiny s kyvadlom:
- malé závažie na kyvadle zodpovedá maternica;
- samotné kyvadlo je vaječníkov, párové pohlavné žľazy ženy;
- vodivá os, na ktorej je pripevnené kyvadlo, je hypotalamus, hlavný "hodinový stroj" menštruačného cyklu;
- kotviaca vidlica, ktorá prenáša pohyby kyvadla na prevody je časť subkortikálnej štruktúry mozgu;
- mechanizmus, ktorý pohybuje ručičkami ciferníka - časť mozgovej kôry, ktorá reguluje hormonálny cyklus.
A závažia alebo pružina naťahovania hodiniek je genetický kód, pokiaľ je to naprogramované, toľko času a celý mechanizmus bude fungovať.
Analogicky s kukučkovými hodinami alebo bojom - to je, ktorých absencia naznačuje poruchu funkcie hodín, to znamená nepravidelnosť menštruačného cyklu.
Kyvadlo, ako viete, sa pohybuje striedavo: najprv v jednom smere, potom v druhom smere, čo zodpovedá dvom fázam menštruačného cyklu.
Nie je potrebné mať povolanie hodinára - každá osoba si bude môcť všimnúť poruchu hodiniek tým, že poruší ich prácu: ponáhľajú sa, zaostávajú, zastavili sa, nevolajú.
Takže ženy môžu určiť stav svojho zdravia jednoduchými znakmi:
Pravidelnosť menštruácie zmizla - porucha. Žiadna ovulácia - nehoda! Absencia menštruácie počas nezvyčajného tehotenstva - katastrofa.
Vlastnosti ženského tela počas rôznych fáz menštruačného cyklu
 Prvá fáza Hormonálny cyklus má za cieľ pripraviť ženu na počatie dieťaťa. To si vyžaduje absolútne zdravé bunky vo všetkých orgánoch a systémoch.
Prvá fáza Hormonálny cyklus má za cieľ pripraviť ženu na počatie dieťaťa. To si vyžaduje absolútne zdravé bunky vo všetkých orgánoch a systémoch.
Preto v tele dominuje sympatický nervový systém, ktorý je regulovaný adrenalínom a noradrenalínom – hormónmi „uteč a bojuj“.
Všetky orgány a systémy ženského tela v tomto období fungujú úplne rovnako. ako v stresovej situácii.
Po ovulácii obraz sa mení. Sada hormonálneho pozadia gestagény sú konzervačné hormóny. Teraz je rast buniek nahradený ich dozrievaním.
Pri regulácii práce orgánov prevláda parasympatický nervový systém, ktorého pôsobenie je zamerané na odstraňovanie následkov stresových situácií.
Praktický význam vedomostí o charakteristikách hormonálneho pozadia v rôznych fázach cyklu
V prvej fáze cyklu užívanie stimulačných liekov bude neúčinné. To platí nielen pre lieky, ktoré zlepšujú pamäť a pozornosť, ale aj pre imunomodulátory.
Telo už pracuje na hranici svojich možností a poháňať ho v prvej fáze je nielen zbytočné, ale ani bezpečné.
Naopak, prostriedky používané na boj proti stresu budú mať najlepší účinok v prvej fáze cyklu, zatiaľ čo v druhom - budú zbytočné.
V druhej fáze- všetko je presne naopak. Zobrazujú sa akékoľvek stimulanty a sedatíva, vrátane trankvilizérov, nemajú požadovaný účinok.
Prečo je potrebné regulovať menštruačný cyklus?
Keď sa ženské telo cyklicky omladzuje, je chránené pred všetkými chorobami staroby, každý kardiológ povie, že na ženy po ukončení cyklickej funkcie číhajú všetky choroby srdcovo-cievneho systému a pred týmto vekom infarkty, resp. hypertenzia je „mužské privilégium“.
Prečo sa celé ľudské telo omladzuje? Na zabezpečenie normálneho života existuje neustály proces nahrádzania niektorých buniek inými, a to aj u mužov. ale v mužskom tele neexistuje jasná „hodinová“ organizácia.
Osvedčenéže nahradenie niektorých buniek inými u žien nastáva všade a každý mesiac. Takže vrstva, ktorá je odmietnutá počas menštruácie, je nahradená v ďalšej fáze cyklu, a to nielen v maternici.
Analýza bunkových zoškrabov z bukálnej sliznice ukazuje rovnaký jav. Tento objav bol urobený ešte v päťdesiatych rokoch 20. storočia.
Teda raz za mesiac pri normálnom cykle dochádza k úplnej výmene buniek v celom tele: z kože do kostnej drene. To je dôvod, prečo je potrebné upraviť menštruačný cyklus pri najmenšej odchýlke.
Korekcia menštruačného cyklu
 Teraz ladíme dvojfázový hormonálny cyklus nepredstavuje žiadne ťažkosti.
Teraz ladíme dvojfázový hormonálny cyklus nepredstavuje žiadne ťažkosti.
Známe hormóny, ktoré sa vyrábajú v prvej a druhej fáze. Syntetizované kombinované lieky ktoré udržujú správnu rovnováhu hormónov.
Ide o bežné antikoncepčné tabletky. Keď sú predpísané na normalizáciu cyklu, antikoncepčný účinok sa považuje za vedľajší účinok.
Výber hormonálneho činidla predpisuje iba lekár. Je potrebné vziať do úvahy nasledujúce parametre:
- spôsobiť, čo spôsobilo porušenie cyklu;
- ústava(individuálne znaky stavby tela) ženy;
- prítomnosť alebo neprítomnosť prejavy mužských sexuálnych charakteristík (rast vlasov na koži, temperament);
- stav mliečnych žliaz a žilový obehový systém.
normálny cyklus je kľúčom k zdraviu ženy. Najmenšie zmeny v menštruačnom cykle podliehajú okamžitej korekcii. Len za takýchto podmienok je možné, aby ženské telo plne fungovalo.
Menštruačný cyklus- cyklicky sa opakujúce zmeny v tele ženy, najmä v častiach reprodukčného systému, ktorých vonkajším prejavom je výtok krvi z pohlavného traktu - menštruácia. Menštruačný cyklus vzniká po menarche (prvej menštruácii) a pretrváva počas celého reprodukčného obdobia života ženy so schopnosťou reprodukovať potomstvo. Cyklické zmeny v tele ženy sú dvojfázové. Prvá (folikulínová) fáza cyklu je určená dozrievaním folikulu a vajíčka vo vaječníku, po ktorom praskne a vajíčko ho opustí – ovulácia. Druhá (luteálna) fáza je spojená s tvorbou žltého telieska.
Súčasne v cyklickom režime dochádza v endometriu postupne k regenerácii a proliferácii funkčnej vrstvy, ktorá je nahradená sekrečnou aktivitou jeho žliaz. Zmeny v endometriu končia deskvamáciou funkčnej vrstvy (menštruácia). Biologický význam zmien, ku ktorým dochádza počas menštruačného cyklu vo vaječníkoch a endometriu, spočíva v zabezpečení reprodukčnej funkcie v štádiách dozrievania vajíčka, jeho oplodnenia a uhniezdenia embrya v maternici. Ak nedôjde k oplodneniu vajíčka, funkčná vrstva endometria sa odmietne, objaví sa krvavý výtok z pohlavného traktu a v reprodukčnom systéme sa znova a v rovnakom poradí vyskytujú procesy zamerané na zabezpečenie dozrievania vajíčka.
Menštruácia- ide o krvavý výtok z pohlavného traktu, ktorý sa v určitých intervaloch opakuje počas celého reprodukčného obdobia života ženy mimo tehotenstva a dojčenia. Menštruácia je vyvrcholením menštruačného cyklu a vyskytuje sa na konci jeho luteálnej fázy v dôsledku odmietnutia funkčnej vrstvy endometria. Prvá menštruácia (menarhe) nastáva vo veku 10-12 rokov. Najbližšieho 1 - 1,5 roka môže byť menštruácia nepravidelná a až potom sa vytvorí pravidelný menštruačný cyklus. Prvý deň menštruácie sa podmienečne považuje za prvý deň cyklu a trvanie cyklu sa vypočíta ako interval medzi prvými dňami dvoch nasledujúcich menštruácií.
menštruačná reprodukčná ovulácia gynekológia
Ryža. jeden. Hormonálna regulácia menštruačného cyklu (schéma): a - mozog; b - zmeny vo vaječníku; c - zmena hladiny hormónov; d - zmeny v endometriu
Vonkajšie parametre normálneho menštruačného cyklu: trvanie od 21 do 35 dní (u 60 % žien je priemerná dĺžka cyklu 28 dní); trvanie menštruačného toku od 2 do 7 dní; množstvo straty krvi v dňoch menštruácie 40-60 ml (priemerne 50 ml).
Procesy, ktoré zabezpečujú normálny priebeh menštruačného cyklu, sú regulované jediným funkčným neuroendokrinným systémom, ktorý zahŕňa centrálne (integrujúce) oddelenia a periférne (efektorové) štruktúry s určitým počtom medzičlánkov. Neuroendokrinnú reguláciu možno v súlade s ich hierarchiou (od vyšších regulačných štruktúr po priame výkonné orgány) rozdeliť do 5 úrovní interagujúcich podľa princípu priamych a inverzných pozitívnych a negatívnych vzťahov (obr.
Prvá (najvyššia) úroveň regulácie fungovanie reprodukčného systému sú štruktúry, ktoré tvoria akceptor všetkých vonkajších a vnútorných (z podriadených oddelení) vplyvov - mozgová kôra centrálneho nervového systému a extrahypotalamické mozgové štruktúry (limbický systém, hipokampus, amygdala). Primeranosť vnímania vonkajších vplyvov CNS a v dôsledku toho aj jeho vplyv na podriadené oddelenia, ktoré regulujú procesy v reprodukčnom systéme, závisia od charakteru vonkajších stimulov (sila, frekvencia a dĺžka ich pôsobenia), ako aj na počiatočnom stave CNS, čo ovplyvňuje jeho odolnosť voči stresovým zaťaženiam.
Je dobre známa možnosť zastavenia menštruácie pri silnom strese (strata blízkych, vojnové podmienky a pod.), ako aj bez zjavných vonkajších vplyvov s celkovou psychickou nerovnováhou („falošné tehotenstvo“ – oneskorenie menštruácie so silnou túžbou alebo so silným strachom otehotnieť). Vyššie regulačné oddelenia reprodukčného systému vnímajú vnútorné vplyvy prostredníctvom špecifických receptorov pre hlavné pohlavné hormóny: estrogény, progesterón a androgény. V reakcii na vonkajšie a vnútorné podnety v mozgovej kôre a extrahypotalamických štruktúrach dochádza k syntéze, uvoľňovaniu a metabolizmu neuropeptidov, neurotransmiterov, ako aj k tvorbe špecifických receptorov, ktoré následne selektívne ovplyvňujú syntézu a uvoľňovanie uvoľňujúceho hormónu hypotalamu. K najdôležitejším neurotransmiterom, t.j. Medzi prenášače patrí norepinefrín, dopamín, kyselina gama-aminomaslová (GABA), acetylcholín, serotonín a melatonín. Mozgové neurotransmitery regulujú produkciu hormónu uvoľňujúceho gonadotropín (GnRH): norepinefrín, acetylcholín a GABA stimulujú ich uvoľňovanie, zatiaľ čo dopamín a serotonín majú opačný účinok.
Neuropeptidy(endogénne opioidné peptidy - EOP, faktor uvoľňujúci kortikotropín a galanín) ovplyvňujú aj funkciu hypotalamu a rovnováhu fungovania všetkých častí reprodukčného systému. V súčasnosti existujú 3 skupiny EOP: enkefalíny, endorfíny a dynorfíny. Tieto látky sa nachádzajú nielen v rôznych štruktúrach mozgu a autonómneho nervového systému, ale aj v pečeni, pľúcach, pankrease a iných orgánoch, ako aj v niektorých biologických tekutinách (krvná plazma, obsah folikulov). Podľa moderných koncepcií sa EOP podieľajú na regulácii tvorby GnRH. Zvýšenie hladiny EOP potláča sekréciu GnRH, a tým aj uvoľňovanie LH a FSH, ktoré môžu byť príčinou anovulácie a v závažnejších prípadoch aj amenorey. Práve s nárastom EOP je výskyt rôznych foriem amenorey centrálnej genézy spojený so stresom, ako aj s nadmernou fyzickou námahou, napríklad u športovcov. Vymenovanie inhibítorov opioidných receptorov (lieky ako naloxón) normalizuje tvorbu GnRH, čo prispieva k normalizácii ovulačnej funkcie a iných procesov v reprodukčnom systéme u pacientov s centrálnou amenoreou. Pri znížení hladiny pohlavných steroidov (s vekom podmieneným alebo chirurgickým odstavením ovariálnej funkcie) nemá EOP inhibičný účinok na uvoľňovanie GnRH, čo pravdepodobne spôsobuje zvýšenú produkciu gonadotropínov u žien po menopauze. Rovnováha syntézy a následných metabolických premien neurotransmiterov, neuropeptidov a neuromodulátorov v mozgových neurónoch a suprahypotalamických štruktúrach teda zabezpečuje normálny priebeh procesov spojených s ovulačnými a menštruačnými funkciami.
Druhá úroveň regulácie reprodukčnou funkciou je hypotalamus, najmä jeho hypofyzárna zóna, pozostávajúca z neurónov ventro- a dorzomediálnych oblúkových jadier, ktoré majú neurosekrečnú aktivitu. Tieto bunky majú vlastnosti neurónov (reprodukujúcich regulačné elektrické impulzy) aj endokrinných buniek, ktoré majú buď stimulačný (liberín) alebo blokujúci (statín) účinok. Aktivita neurosekrécie v hypotalame je regulovaná pohlavnými hormónmi, ktoré prichádzajú z krvného obehu, ako aj neurotransmitermi a neuropeptidmi tvorenými v mozgovej kôre a suprahypotalamických štruktúrach. Hypotalamus vylučuje GnRH, ktorý obsahuje folikuly stimulujúce (RGFSH – folliberín) a luteinizačné (RGLG – luliberín) hormóny, ktoré pôsobia na hypofýzu. Uvoľňujúci hormón LH (RGLG - luliberín) bol izolovaný, syntetizovaný a podrobne opísaný. Doposiaľ nebolo možné izolovať a syntetizovať hormón stimulujúci uvoľňovanie folikulov. Zistilo sa však, že dekapeptid RGLG a jeho syntetické analógy stimulujú uvoľňovanie pohlavných žliaz prostredníctvom trofov nielen LH, ale aj FSH. V tejto súvislosti sa pre gonadotropné liberíny ujal jeden termín – gonadotropín uvoľňujúci hormón (GnRH), ktorý je v podstate synonymom pre RHLH. Hypotalamický liberín, ktorý stimuluje tvorbu prolaktínu, tiež nebol identifikovaný, hoci sa zistilo, že jeho syntézu aktivuje hormón uvoľňujúci TSH (tyroliberín). Tvorba prolaktínu je tiež aktivovaná serotonínom a endogénnymi opioidnými peptidmi, ktoré stimulujú serotonergné systémy. Dopamín, naopak, inhibuje uvoľňovanie prolaktínu z laktotrofov adenohypofýzy. Použitie dopaminergných liekov, ako je parlodel (bromkriptín), môže úspešne liečiť funkčnú a organickú hyperprolaktinémiu, ktorá je veľmi častou príčinou porúch menštruácie a ovulácie. Sekrécia GnRH je geneticky naprogramovaná a má pulzačný (cirhorálny) charakter: vrcholy zvýšenej sekrécie hormónov trvajúce niekoľko minút sú nahradené 1-3-hodinovými intervalmi relatívne nízkej sekrečnej aktivity. Frekvencia a amplitúda sekrécie GnRH reguluje hladinu estradiolu – emisie GnRH v predovulačnej perióde na pozadí maximálneho uvoľňovania estradiolu sú výrazne väčšie ako v skorých folikulárnych a luteálnych fázach.
Tretia úroveň regulácie reprodukčná funkcia je predná hypofýza, v ktorej sa vylučujú gonadotropné hormóny – folikuly stimulujúce, čiže folitropín (FSH), a luteinizačné, čiže lutropín (LH), prolaktín, adrenokortikotropný hormón (ACTH), somatotropný hormón (STH) a štítna žľaza- stimulačný hormón (TSH). Normálne fungovanie reprodukčného systému je možné len pri vyváženom výbere každého z nich. FSH stimuluje rast a dozrievanie folikulov vo vaječníku, proliferáciu granulóznych buniek; tvorba FSH a LH receptorov na granulózových bunkách; aktivita aromatázy v dozrievajúcom folikule (to zvyšuje konverziu androgénov na estrogény); produkciu inhibínu, aktivínu a inzulínu podobných rastových faktorov. LH podporuje tvorbu androgénov v bunkách theca; ovulácia (spolu s FSH); prestavba granulóznych buniek počas luteinizácie; syntéza progesterónu v corpus luteum. Prolaktín má rôzne účinky na telo ženy. Jeho hlavnou biologickou úlohou je stimulovať rast mliečnych žliaz, regulovať laktáciu a tiež kontrolovať sekréciu progesterónu žltým telieskom aktiváciou tvorby receptorov pre LH v ňom.V tehotenstve a počas laktácie prestáva inhibícia syntézy prolaktínu a , v dôsledku toho zvýšenie jeho hladiny v krvi .
Do štvrtej úrovne regulácia reprodukčnej funkcie zahŕňajú periférne endokrinné orgány (vaječníky, nadobličky, štítna žľaza). Hlavná úloha patrí vaječníkom a ostatné žľazy vykonávajú svoje špecifické funkcie pri zachovaní normálneho fungovania reprodukčného systému. Vo vaječníkoch dochádza k rastu a dozrievaniu folikulov, ovulácii, tvorbe žltého telieska a syntéze pohlavných steroidov. Pri narodení obsahujú vaječníky dievčaťa približne 2 milióny primordiálnych folikulov. Väčšina z nich prechádza počas života atretickými zmenami a len veľmi malá časť prejde úplným vývojovým cyklom od prvotného po zrelý s následným vytvorením žltého telieska. V čase menarche obsahujú vaječníky 200-400 tisíc prvotných folikulov. Počas jedného menštruačného cyklu sa spravidla vyvinie iba jeden folikul s vajíčkom vo vnútri. V prípade dozrievania väčšieho počtu je možná viacplodová gravidita.
Folikulogenéza začína vplyvom FSH v neskorej časti luteálnej fázy cyklu a končí na začiatku vrcholu uvoľňovania gonadotropínu. Približne 1 deň pred nástupom menštruácie hladina FSH opäť stúpa, čo zabezpečuje vstup do rastu, resp. nábor folikulov (1--4. deň cyklu), výber folikulu z kohorty homogénnych - kvázi -synchronizované (5--7. deň), dozrievanie dominantného folikulu (8-12. deň) a ovulácia (13.-15. deň). Tento proces, ktorý tvorí folikulárnu fázu, trvá približne 14 dní. V dôsledku toho sa vytvorí predovulačný folikul a zvyšok kohorty folikulov, ktoré vstúpili do rastu, podlieha atrézii. Výber jediného folikulu určeného na ovuláciu je neoddeliteľný od syntézy estrogénu v ňom. Stabilita produkcie estrogénu závisí od interakcie medzi bunkami theca a granulóznymi bunkami, ktorých aktivita je zasa modulovaná početnými endokrinnými, parakrinnými a autokrinnými mechanizmami, ktoré regulujú rast a dozrievanie folikulov. V závislosti od štádia vývoja a morfologických znakov sa rozlišujú primordiálne, preantrálne, antrálne a preovulačné alebo dominantné folikuly. Primordiálny folikul pozostáva z nezrelého vajíčka, ktoré sa nachádza vo folikulárnom a granulárnom (granulárnom) epiteli. Vonku je folikul obklopený membránou spojivového tkaniva (bunky theca). Počas každého menštruačného cyklu začne rásť 3 až 30 primordiálnych folikulov, ktoré sa transformujú na preantrálne (primárne) folikuly. preantrálny folikul. V preantrálnom folikule sa oocyt zväčšuje a je obklopený membránou nazývanou zona pellucida. Bunky granulózneho epitelu proliferujú a zaokrúhľujú sa, čím vytvárajú zrnitú vrstvu folikulu (stratum granulosum) a z okolitej strómy sa vytvára vrstva theca. Toto štádium je charakterizované aktiváciou produkcie estrogénov tvorených v granulóznej vrstve.
Preovulačný (dominantný) folikul(obr. 2.2) vyniká medzi rastúcimi folikulmi najväčšou veľkosťou (priemer v čase ovulácie dosahuje 20 mm). Dominantný folikul má bohato vaskularizovanú vrstvu buniek theca a buniek granulózy s veľkým počtom receptorov pre FSH a LH. Spolu s rastom a vývojom dominantného predovulačného folikulu vo vaječníkoch sa paralelne vyskytuje atrézia zvyšných (rekrutovaných) folikulov, ktoré pôvodne vstúpili do rastu, a pokračuje aj atrézia primordiálnych folikulov. Počas dozrievania dochádza k 100-násobnému zväčšeniu objemu folikulárnej tekutiny v preovulačnom folikule. V procese dozrievania antrálnych folikulov sa mení zloženie folikulárnej tekutiny.
Antrálny (sekundárny) folikul prechádza zväčšením dutiny vytvorenej akumulujúcou sa folikulárnou tekutinou produkovanou bunkami granulóznej vrstvy. Zvyšuje sa aj aktivita tvorby pohlavných steroidov. Bunky theca syntetizujú androgény (androstendión a testosterón). Keď sú androgény v granulózových bunkách aktívne aromatizované, čo určuje ich premenu na estrogény. Vo všetkých štádiách vývoja folikulov, okrem predovulačného, je obsah progesterónu na konštantnej a relatívne nízkej úrovni. Gonadotropínov a prolaktínu je vo folikulárnej tekutine vždy menej ako v krvnej plazme a hladina prolaktínu má tendenciu klesať, keď folikul dozrieva. FSH sa určuje od začiatku tvorby kavity a LH možno detegovať len v zrelom predovulačnej folikule spolu s progesterónom. Folikulárna tekutina obsahuje aj oxytocín a vazopresín a v koncentráciách 30-krát vyšších ako v krvi, čo môže naznačovať lokálnu tvorbu týchto neuropeptidov. Prostaglandíny triedy E a F sa detegujú iba v predovulačných folikuloch a až po začatí vzostupu hladiny LH, čo naznačuje ich priame zapojenie do procesu ovulácie.
Ovulácia- prasknutie preovulačného (dominantného) folikulu a uvoľnenie vajíčka z neho. Ovuláciu sprevádza krvácanie zo zničených kapilár obklopujúcich bunky théky (obr. 2.3). Predpokladá sa, že ovulácia nastáva 24-36 hodín po predovulačnej špičke estradiolu, čo spôsobuje prudké zvýšenie sekrécie LH. Na tomto pozadí sa aktivujú proteolytické enzýmy – kolagenáza a plazmín, ktoré ničia kolagén steny folikulu a tým znižujú jeho pevnosť. Zároveň pozorované zvýšenie koncentrácie prostaglandínu F2a, ako aj oxytocínu, vyvoláva ruptúru folikulu v dôsledku ich stimulácie kontrakcie hladkého svalstva a vypudenia oocytu s vajcovodným valčekom z dutiny folikulu. . Prasknutie folikulu je tiež uľahčené zvýšením koncentrácie prostaglandínu E2 a relaxínu v ňom, ktoré znižujú tuhosť jeho stien. Po uvoľnení vajíčka vzniknuté kapiláry rýchlo prerastú do dutiny ovulovaného folikulu. Bunky granulózy prechádzajú luteinizáciou, morfologicky sa prejavujúcou zväčšením ich objemu a tvorbou lipidových inklúzií. Tento proces, ktorý vedie k vytvoreniu žltého telieska, je stimulovaný LH, ktorý aktívne interaguje so špecifickými receptormi granulóznych buniek.
corpus luteum- prechodný hormonálne aktívny útvar, fungujúci 14 dní bez ohľadu na celkové trvanie menštruačného cyklu. Ak nedôjde k otehotneniu, dochádza k regresii corpus luteum. Úplné žlté teliesko vzniká až vo fáze, keď sa v preovulačnom folikule vytvorí primerané množstvo granulóznych buniek s vysokým obsahom LH receptorov.V reprodukčnom období sú vaječníky hlavným zdrojom estrogénov (estradiol, estriol a estrón) , z ktorých je najaktívnejší estradiol. Okrem estrogénov sa vo vaječníkoch tvorí progesterón a určité množstvo androgénov. Okrem steroidných hormónov a inhibínov, ktoré sa dostávajú do krvného obehu a ovplyvňujú cieľové orgány, sa vo vaječníkoch syntetizujú aj biologicky aktívne zlúčeniny s prevažne lokálnym účinkom podobným hormónom. Vytvorené prostaglandíny, oxytocín a vazopresín teda hrajú dôležitú úlohu ako spúšťače ovulácie. Oxytocín má tiež luteolytický účinok, poskytuje regresiu žltého telieska. Relaxín podporuje ovuláciu a má tokolytický účinok na myometrium. Rastové faktory – epidermálny rastový faktor (EGF) a inzulínu podobné rastové faktory 1 a 2 (IPGF-1 a IPFR-2) aktivujú proliferáciu granulóznych buniek a dozrievanie folikulov. Rovnaké faktory sa spolu s gonadotropínmi podieľajú na jemnej regulácii procesov selekcie dominantného folikulu, atrézie degenerujúcich folikulov všetkých štádií, ako aj na ukončení fungovania žltého telieska. Tvorba androgénov vo vaječníkoch zostáva stabilná počas celého cyklu. Hlavným biologickým účelom cyklickej sekrécie pohlavných steroidov vo vaječníku je regulácia fyziologických cyklických zmien v endometriu. Ovariálne hormóny neurčujú len funkčné zmeny v samotnom reprodukčnom systéme. Aktívne ovplyvňujú aj metabolické procesy v iných orgánoch a tkanivách, ktoré majú receptory pre pohlavné steroidy. Tieto receptory môžu byť buď cytoplazmatické (cytosolové receptory) alebo jadrové.
Cytoplazmatické receptory sú prísne špecifické pre estrogén, progesterón a testosterón, zatiaľ čo jadrové receptory môžu prijímať nielen steroidné hormóny, ale aj aminopeptidy, inzulín a glukagón. Pre väzbu na progesterónový receptor sa glukokortikoidy považujú za antagonisty. V pokožke sa vplyvom estradiolu a testosterónu aktivuje syntéza kolagénu, ktorá pomáha udržiavať jej elasticitu. Zvýšený kožný maz, akné, folikulitída, pórovitosť a nadmerné ochlpenie sú spojené so zvýšenou expozíciou androgénom. V kostiach estrogény, progesterón a androgény podporujú normálnu prestavbu tým, že bránia kostnej resorpcii. V tukovom tkanive rovnováha estrogénov a androgénov predurčuje tak aktivitu jeho metabolizmu, ako aj distribúciu v organizme. Pohlavné steroidy (progesterón) významne modulujú prácu hypotalamického termoregulačného centra. S receptormi pre pohlavné steroidy v centrálnom nervovom systéme, v štruktúrach hipokampu, ktoré regulujú emocionálnu sféru, ako aj v centrách, ktoré riadia autonómne funkcie, sa spája fenomén „menštruačnej vlny“ v dňoch pred menštruáciou. Tento jav sa prejavuje nerovnováhou v procesoch aktivácie a inhibície v kôre, kolísaním tonusu sympatického a parasympatického systému (zvlášť výrazne ovplyvňujúcim fungovanie kardiovaskulárneho systému), ako aj zmenami nálady a určitou podráždenosťou. U zdravých žien však tieto zmeny neprekračujú fyziologické hranice.
Piata úroveň regulácia reprodukčných funkcií sú citlivé na kolísanie hladín pohlavných steroidov vnútorné a vonkajšie časti reprodukčného systému (maternica, vajíčkovody, pošvová sliznica), ako aj mliečne žľazy. Najvýraznejšie cyklické zmeny sa vyskytujú v endometriu.
Cyklické zmeny v endometriu dotýkať sa jeho povrchovej vrstvy, pozostávajúcej z kompaktných epiteliálnych buniek, a medziproduktu, ktoré sú počas menštruácie odmietnuté. Bazálna vrstva, ktorá sa počas menštruácie neodmieta, zabezpečuje obnovu deskvamovaných vrstiev. Podľa zmien endometria počas cyklu sa rozlišuje proliferačná fáza, sekréčná fáza a fáza krvácania (menštruácia).
Proliferačná fáza(„folikulárny") trvá v priemere 12-14 dní, počnúc 5. dňom cyklu. V tomto období sa vytvára nová povrchová vrstva s predĺženými tubulárnymi žľazami lemovanými cylindrickým epitelom so zvýšenou mitotickou aktivitou. Hrúbka tzv. funkčná vrstva endometria je 8 mm.
Fáza sekrécie (luteálna) spojená s činnosťou žltého telieska, trvá 14 dní (+ 1 deň). Počas tohto obdobia začne epitel endometriálnych žliaz produkovať tajomstvo obsahujúce kyslé glykozaminoglykány, glykoproteíny a glykogén. Aktivita sekrécie sa stáva najvyššou na 20-21 deň. Do tejto doby sa v endometriu nachádza maximálne množstvo proteolytických enzýmov a v stróme nastávajú deciduálne transformácie (bunky kompaktnej vrstvy sa zväčšujú, nadobúdajú zaoblený alebo polygonálny tvar, v ich cytoplazme sa hromadí glykogén). Dochádza k prudkej vaskularizácii strómy – špirálové tepny sú ostro kľukaté, tvoria „spletence“ nachádzajúce sa v celej funkčnej vrstve. Žily sú rozšírené. Takéto zmeny v endometriu, pozorované na 20. až 22. deň (6. až 8. deň po ovulácii) 28-dňového menštruačného cyklu, poskytujú najlepšie podmienky pre implantáciu oplodneného vajíčka. Do 24.-27. dňa je v dôsledku začiatku regresie žltého telieska a poklesu koncentrácie ním produkovaných hormónov narušený trofizmus endometria s postupným zvyšovaním degeneratívnych zmien v ňom. Z granulárnych buniek endometriálnej strómy sa uvoľňujú granuly obsahujúce relaxín, ktorý pripravuje menštruačné odmietnutie sliznice. V povrchových oblastiach kompaktnej vrstvy je zaznamenaná lacunárna expanzia kapilár a krvácanie v stróme, ktoré je možné zistiť za 1 deň. pred nástupom menštruácie.
Menštruácia zahŕňa deskvamáciu a regeneráciu funkčnej vrstvy endometria. V dôsledku regresie žltého telieska a prudkého poklesu obsahu pohlavných steroidov v endometriu sa zvyšuje hypoxia. Nástup menštruácie je uľahčený predĺženým kŕčom tepien, čo vedie k stagnácii krvi a tvorbe krvných zrazenín. Tkanivovú hypoxiu (tkanivovú acidózu) zhoršuje zvýšená permeabilita endotelu, krehkosť cievnych stien, početné malé krvácania a masívna infiltrácia leukocytmi. Lysozomálne proteolytické enzýmy uvoľňované z leukocytov zvyšujú topenie tkanivových prvkov. Po dlhotrvajúcom spazme ciev dochádza k ich paretickej expanzii so zvýšeným prietokom krvi. Súčasne sa zaznamená zvýšenie hydrostatického tlaku v mikrovaskulatúre a prasknutie stien ciev, ktoré už do značnej miery stratili svoju mechanickú pevnosť. Na tomto pozadí dochádza k aktívnej deskvamácii nekrotických oblastí funkčnej vrstvy. Do konca 1. dňa menštruácie sú odmietnuté 2/3 funkčnej vrstvy a jej úplná deskvamácia sa zvyčajne končí na 3. deň. Regenerácia endometria začína ihneď po odmietnutí nekrotickej funkčnej vrstvy.
Základom pre regeneráciu sú epitelové bunky strómy bazálnej vrstvy. Za fyziologických podmienok už na 4. deň cyklu dochádza k epitelizácii celého povrchu rany na sliznici. Nasledujú opäť cyklické zmeny v endometriu – fázy proliferácie a sekrécie. Postupné zmeny počas cyklu v endometriu: proliferácia, sekrécia a menštruácia závisia nielen od cyklických výkyvov hladín pohlavných steroidov v krvi, ale aj od stavu tkanivových receptorov pre tieto hormóny. Koncentrácia jadrových estradiolových receptorov sa zvyšuje až do polovice cyklu, pričom vrchol dosahuje v neskoršom období fázy proliferácie endometria. Po ovulácii dochádza k rýchlemu poklesu koncentrácie jadrových estradiolových receptorov, ktorý pokračuje až do neskorej sekrečnej fázy, kedy je ich expresia výrazne nižšia ako na začiatku cyklu. Zistilo sa, že indukcia tvorby receptorov pre estradiol aj progesterón závisí od koncentrácie estradiolu v tkanivách. Vo včasnej proliferačnej fáze je obsah progesterónových receptorov nižší ako obsah estradiolu, potom však dochádza k predovulačnému vzostupu hladiny progesterónových receptorov.
Po ovulácii dosiahne hladina jadrových receptorov pre progesterón maximum za celý cyklus. V proliferatívnej fáze estradiol priamo stimuluje tvorbu progesterónových receptorov, čo vysvetľuje nedostatok vzťahu medzi plazmatickými hladinami progesterónu a obsahom jeho receptorov v endometriu. Regulácia lokálnej koncentrácie estradiolu a progesterónu je do značnej miery sprostredkovaná výskytom rôznych enzýmov počas menštruačného cyklu. Obsah estrogénov v endometriu závisí nielen od ich hladiny v krvi, ale aj od vzdelania. Endometrium ženy je schopné syntetizovať estrogény premenou androstendiónu a testosterónu za účasti aromatázy (aromatizácia). Tento lokálny zdroj estrogénu zvyšuje estrogenizáciu endometriálnych buniek, ktorá charakterizuje proliferatívnu fázu. Počas tejto fázy je zaznamenaná najvyššia aromatizácia androgénov a najnižšia aktivita enzýmov metabolizujúcich estrogén. Nedávno sa zistilo, že endometrium je schopné vylučovať prolaktín, ktorý je úplne identický s hypofýzou. Syntéza prolaktínu endometriom začína v druhej polovici luteálnej fázy (aktivovaná progesterónom) a zhoduje sa s decidualizáciou stromálnych buniek. Cyklická aktivita reprodukčného systému je určená princípmi priamej a spätnej väzby, ktorú zabezpečujú špecifické hormonálne receptory v každom z väzieb. Priamou väzbou je stimulačný účinok hypotalamu na hypofýzu a následná tvorba pohlavných steroidov vo vaječníku. Spätná väzba je určená vplyvom zvýšenej koncentrácie pohlavných steroidov na nadložné hladiny. Pri interakcii väzieb reprodukčného systému sa rozlišujú „dlhé“, „krátke“ a „ultrakrátke“ slučky. "Dlhá" slučka je účinok cez receptory hypotalamo-hypofyzárneho systému na produkciu pohlavných hormónov. "Krátka" slučka definuje spojenie medzi hypofýzou a hypotalamom. "Ultrakrátka" slučka je spojenie medzi hypotalamom a nervovými bunkami, ktoré vykonávajú lokálnu reguláciu pomocou neurotransmiterov, neuropeptidov, neuromodulátorov a elektrických stimulov.
Posúdenie stavu reprodukčného systému podľa testov funkčnej diagnostiky. Už dlhé roky sa v gynekologickej praxi používajú takzvané testy funkčnej diagnostiky stavu reprodukčného systému. Hodnota týchto pomerne jednoduchých štúdií sa zachovala až do súčasnosti. Najčastejšie sa využíva meranie bazálnej teploty, hodnotenie fenoménu „zornice“ a hlienu krčka maternice (kryštalizácia, distenzibilita), ako aj výpočet karyopyknotického indexu (KPI, %) pošvového epitelu.
Test bazálnej teploty je založená na schopnosti progesterónu (vo zvýšenej koncentrácii) spôsobiť reštrukturalizáciu práce centra termoregulácie hypotalamu, čo vedie k prechodnej hypertermickej reakcii. Teplota sa meria denne v konečníku ráno bez toho, aby ste vstali z postele. Výsledky sú zobrazené graficky. Pri normálnom dvojfázovom menštruačnom cykle stúpa bazálna teplota v progesterónovej fáze o 0,4 - 0,8 ° C. V deň menštruácie alebo 1 deň pred jej začiatkom sa bazálna teplota znižuje. Pretrvávajúci dvojfázový cyklus (bazálna teplota by sa mala merať počas 2-3 menštruačných cyklov) naznačuje, že nastala ovulácia a funkčne aktívne žlté teliesko. Neprítomnosť zvýšenia teploty v druhej fáze cyklu naznačuje anovuláciu, oneskorenie vzostupu a / alebo jeho krátke trvanie (zvýšenie teploty o 2--7 dní) - skrátenie luteálnej fázy, nedostatočný vzostup (o 0,2-0,3 °C) - - pre nedostatočnú funkciu žltého telieska. Falošne pozitívny výsledok (zvýšenie bazálnej teploty pri absencii žltého telieska) môže byť s akútnymi a chronickými infekciami, s niektorými zmenami v centrálnom nervovom systéme, sprevádzanými zvýšenou excitabilitou. Symptóm "žiak" odráža množstvo a stav slizničnej sekrécie v cervikálnom kanáli, ktoré závisia od saturácie tela estrogénmi. Najväčšie množstvo cervikálneho hlienu sa tvorí počas ovulácie, najmenšie - pred menštruáciou. Fenomén "žiačky" je založený na rozšírení vonkajšieho os cervikálneho kanála v dôsledku nahromadenia priehľadného sklovcového hlienu v krčku maternice. V predovulačných dňoch sa rozšírený vonkajší otvor cervikálneho kanála podobá zrenici. Fenomén „žiak“ sa v závislosti od jeho závažnosti odhaduje na 1-3 plusy. Test nemožno použiť na patologické zmeny na krčku maternice.
Posúdenie kvality hlienu krčka maternice odráža jeho kryštalizáciu a stupeň napätia. Kryštalizácia (fenomén papradie) cervikálneho hlienu počas sušenia je najvýraznejšia počas ovulácie, potom sa postupne znižuje a úplne chýba pred menštruáciou. Bodovo (od 1 do 3) sa hodnotí aj kryštalizácia na vzduchu vysušeného hlienu. Napätie hlienu krčka maternice závisí od saturácie estrogénom. Hlien sa odstráni z cervikálneho kanála pomocou klieští, čeľuste nástroja sa oddelia, čím sa určí stupeň napätia. Pred menštruáciou je dĺžka vlákna maximálna (12 cm). Hlien môže byť negatívne ovplyvnený zápalovými procesmi v pohlavných orgánoch, ako aj hormonálnou nerovnováhou.
Karyopyknotický index. Cyklické fluktuácie ovariálnych hormónov sú spojené so zmenami v bunkovom zložení sliznice endometria. V nátere z vagíny sa podľa morfologických znakov rozlišujú 4 typy buniek dlaždicového vrstveného epitelu:
- a) keratinizácia;
- b) medziprodukt;
- c) parabazálne;
- d) bazálny. Karyopyknotický index (KPI) je pomer počtu buniek s pyknotickým jadrom (t.j. keratinizujúcich buniek) k celkovému počtu epitelových buniek v nátere, vyjadrený v percentách.
Vo folikulárnej fáze menštruačného cyklu je CPI 20–40 %, v predovulačných dňoch stúpa na 80–88 % a v luteálnej fáze cyklu klesá na 20–25 %. Kvantitatívne pomery bunkových elementov v náteroch vaginálnej sliznice teda umožňujú posúdiť saturáciu tela estrogénmi.
