Регенерація як властивість живого: здатність до самооновлення та відновлення. Типи регенерації
РЕГЕНЕРАЦІЯ
відновлення організмом втрачених частин тієї чи іншої стадії життєвого циклу. Регенерація зазвичай відбувається у разі ушкодження чи втрати якогось органу чи частини організму. Однак, крім цього, в кожному організмі протягом усього його життя постійно йдуть процеси відновлення та оновлення. Людина, наприклад, постійно оновлюється зовнішній шар шкіри. Птахи періодично скидають пір'я і відрощують нові, а ссавці змінюють шерстий покрив. У листопадних дерев листя щорічно опадає і замінюється свіжим. Таку регенерацію, зазвичай не пов'язану з ушкодженнями чи втратою, називають фізіологічною. Регенерацію, що відбувається після пошкодження чи втрати будь-якої частини тіла, називають репаративною. Тут ми розглянемо лише репаративну регенерацію. Репаративна регенерація може бути типовою або атиповою. При типовій регенерації втрачена частина заміщається шляхом розвитку точно такої ж частини. Причиною втрати може бути зовнішній вплив (наприклад, ампутація), або ж тварина навмисно відриває частину свого тіла (аутотомія), як ящірка, що обламує частину свого хвоста, рятуючись від ворога. При атиповій регенерації втрачена частина заміщається структурою, що відрізняється від початкової кількісно чи якісно. У кінцівки, що регенерувала пуголовка, число пальців може виявитися менше вихідного, а у креветки замість ампутованого ока може вирости антена.
РЕГЕНЕРАЦІЯ У ТВАРИН
Здатність до регенерації поширена серед тварин. Взагалі, нижчі тварини частіше здатні до регенерації, ніж складніші високоорганізовані форми. Так, серед безхребетних набагато більше видів, здатних відновлювати втрачені органи, ніж серед хребетних, але тільки в деяких із них можлива регенерація цілої особини з її невеликого фрагмента. Проте загальне правило зниження здатності до регенерації з підвищенням складності організму не можна вважати абсолютним. Такі примітивні тварини, як гребневики та коловратки, практично не здатні до регенерації, а у набагато складніших ракоподібних та амфібій ця здатність добре виражена; відомі та інші винятки. Деякі близькі споріднені тварини сильно різняться в цьому відношенні. Так, у дощового черв'яка з невеликого шматочка тіла може повністю регенерувати нова особина, тоді як п'явки не здатні відновити один втрачений орган. У хвостатих амфібій на місці ампутованої кінцівки утворюється нова, а у жаби куксу просто гоїться і ніякого нового зростання не відбувається. Багато безхребетних здатні до регенерації значної частини тіла. У губок, гідроїдних поліпів, плоских, стрічкових та кільчастих черв'яків, мошанок, голкошкірих та оболонників з невеликого фрагмента тіла може регенерувати цілий організм. Особливо примітна здатність до регенерації у губок. Якщо тіло дорослої губки продавити через сітчасту тканину, всі клітини відокремляться одна від одної, як просіяні крізь сито. Якщо потім помістити всі ці окремі клітини у воду і обережно, ретельно перемішати, повністю зруйнувавши всі зв'язки між ними, то через деякий час вони починають поступово зближуватися і з'єднуються, утворюючи цілу губку, подібну до колишньої. У цьому бере участь свого роду "впізнавання" на клітинному рівні, про що свідчить наступний експеримент. Губки трьох різних видів розділяли описаним способом на окремі клітини і добре перемішували. При цьому виявилося, що клітини кожного виду здатні "пізнавати" в загальній масі клітини свого виду і возз'єднуються тільки з ними, так що в результаті утворилася не одна, а три нові губки, подібні до трьох вихідних.
Стрічковий черв'як, довжина якого багато разів перевищує його ширину, здатний відтворити цілу особину з будь-якої ділянки свого тіла. Теоретично можливо, розрізавши одного хробака на 200 000 шматочків, отримати з нього в результаті регенерації 200 000 нових хробаків. З одного променя морської зірки може регенерувати ціла зірка.

Молюски, членистоногі та хребетні не здатні регенерувати цілу особину з одного фрагмента, однак у багатьох із них відбувається відновлення втраченого органу. Деякі в разі потреби вдаються до аутотомії. Птахи та ссавці як еволюційно найбільш просунуті тварини менше за інших здатні до регенерації. У птахів можливе заміщення пір'я та деяких частин дзьоба. Ссавці можуть відновлювати покрив, пазурі та частково печінку; вони здатні також до загоєння ран, а олені - до відрощування нових рогів замість скинутих.
Процеси регенерації. У регенерації у тварин беруть участь два процеси: епіморфоз та морфалаксис. При епіморфічній регенерації втрачена частина тіла відновлюється за рахунок активності недиференційованих клітин. Ці клітини, схожі на ембріональні, накопичуються під пораненим епідермісом біля поверхні розрізу, де вони утворюють зачаток або бластему. Клітини бластеми поступово розмножуються і перетворюються на тканини нового органа чи частини тіла. При морфалаксисі інші тканини тіла або органу безпосередньо перетворюються на структури недостатньої частини. У гідроїдних поліпів регенерація відбувається головним чином шляхом морфалаксису, а у планарій у ній одночасно беруть участь і епіморфоз, і морфалаксис. Регенерація шляхом утворення бластеми широко поширена у безхребетних і відіграє важливу роль у регенерації органів у амфібій. Існує дві теорії походження бластемних клітин: 1) клітини бластеми походять із "резервних клітин", тобто. клітин, що залишилися невикористаними в процесі ембріонального розвитку та розподілилися по різних органах тіла; 2) тканини, цілісність яких було порушено при ампутації, " дедиференціюються " у сфері розрізу, тобто. дезінтегруються і перетворюються на окремі бластемні клітини. Таким чином, згідно з теорією "резервних клітин", бластема утворюється з клітин, що залишалися ембріональними, які мігрують з різних ділянок тіла і накопичуються біля поверхні розрізу, а згідно з теорією "дедиференційованої тканини", бластемні клітини походять з клітин пошкоджених тканин. На підтвердження як однієї, і іншої теорії є достатньо даних. Наприклад, у планарій резервні клітини чутливіші до рентгенівських променів, ніж клітини диференційованої тканини; тому їх можна зруйнувати, суворо дозуючи опромінення, ніж пошкодити нормальні тканини планарии. Опромінені таким чином особини виживають, але втрачають здатність до регенерації. Однак якщо тільки передню половину тіла планарії піддати опроміненню, а потім розрізати, то регенерація відбувається хоча і з деякою затримкою. Затримка свідчить про те, що бластема утворюється із резервних клітин, які мігрують на поверхню розрізу з неопроміненої половини тіла. Міграцію цих резервних клітин з опроміненої частини тіла можна спостерігати під мікроскопом. Подібні експерименти показали, що з тритона регенерація кінцівок відбувається з допомогою бластемних клітин місцевого походження, тобто. за рахунок дедиференціювання пошкоджених тканин кукси. Якщо, наприклад, опромінити всю личинку тритону, за винятком, скажімо, правої передньої кінцівки, а потім ампутувати цю кінцівку на рівні передпліччя, то у тварини відростає нова передня кінцівка. Очевидно, що необхідні для цього бластемні клітини надходять саме з кукси передньої кінцівки, так як все інше тіло зазнало опромінення. Більш того, регенерація відбувається навіть у тому випадку, якщо опромінюють всю личинку, за винятком ділянки шириною 1 мм на правій передній лапці, а потім останню ампутують, роблячи розріз через цю неопромінену ділянку. У цьому випадку цілком очевидно, що бластемні клітини надходять із поверхні розрізу, оскільки все тіло, включаючи праву передню лапку, було позбавлене здатності до регенерації. Описані процеси аналізували із застосуванням сучасних методів. Електронний мікроскоп дозволяє спостерігати зміни у пошкоджених та регенеруючих тканинах у всіх деталях. Створені барвники, що виявляють певні хімічні речовини, що містяться у клітинах та тканинах. Гістохімічні методи (із застосуванням барвників) дозволяють судити про біохімічні процеси, що відбуваються при регенерації органів і тканин.
Полярність. Одна з найзагадковіших проблем у біології – походження полярності в організмів. З кулястого яйця жаби розвивається пуголовок, у якого з самого початку на одному кінці тіла знаходиться голова з головним мозком, очима та ротом, а на іншому – хвіст. Так само, якщо розрізати тіло планарії деякі фрагменти, одному кінці кожного фрагмента розвивається голова, але в інший - хвіст. У цьому голова завжди утворюється передньому кінці фрагмента. Експерименти ясно показують, що у планарії існує градієнт метаболічної (біохімічної) активності, що проходить передньо-задньої осі її тіла; при цьому найвищу активність має передній кінець тіла, а в напрямку до заднього кінця активність поступово знижується. У будь-якої тварини голова завжди утворюється на тому кінці фрагмента, де метаболічна активність вища. Якщо напрям градієнта метаболічної активності в ізольованому фрагменті планарії змінити протилежне, те й формування голови відбудеться протилежному кінці фрагмента. Градієнт метаболічної активності в тілі планарій відображає існування якогось важливішого фізико-хімічного градієнта, природа якого поки що невідома. У регенеруючій кінцівці тритону полярність новоутворюваної структури, мабуть, визначається культей, що збереглася. З причин, які ще залишаються незрозумілими, в регенеруючому органі формуються тільки структури, розташовані дистальніше за ранову поверхню, а ті, що розташовані проксимальніше (ближче до тіла), не регенерують ніколи. Так, якщо ампутувати кисть тритону, а частину передньої кінцівки, що залишилася, вставити обрізаним кінцем у стінку тіла і дати цьому дистальному (віддаленому від тіла) кінці прижитися на новому, незвичайному для нього місці, то наступна перерізка цієї верхньої кінцівки поблизу плеча (що звільняє її від зв'язку) з плечем) призводить до регенерації кінцівки з повним набором дистальних структур. У такої кінцівки є на момент перерізання наступні частини (починаючи з зап'ястя, що злився зі стінкою тіла): зап'ястя, передпліччя, лікоть і половина дистальна плеча; потім, в результаті регенерації, з'являються ще одна дистальна половина плеча, лікоть, передпліччя, зап'ястя і кисть. Таким чином, інвертована (перевернута) кінцівка регенерувала всі частини, розташовані дистальніше за ранову поверхню. Це разюче явище вказує на те, що тканини кукси (в даному випадку кукси кінцівки) контролюють регенерацію органу. Завдання подальших досліджень - з'ясувати, які саме фактори контролюють цей процес, що стимулює регенерацію і що змушує клітини, що забезпечують регенерацію, накопичуватися на поверхні рани. Деякі вчені вважають, що пошкоджені тканини виділяють якийсь хімічний "рановий фактор". Однак виділити хімічну речовину, специфічну для ран, поки що не вдалося.
РЕГЕНЕРАЦІЯ У РОСЛИН
Широке поширення регенерації в царстві рослин обумовлено збереженням у них меристем (тканин, що складаються з клітин, що діляться) і недиференційованих тканин. Найчастіше регенерація в рослин - це, по суті, одна з форм вегетативного розмноження. Так, на кінчику нормального стебла є верхівкова нирка, що забезпечує безперервне утворення нового листя і зростання стебла в довжину протягом усього життя цієї рослини. Якщо відрізати цю нирку і підтримувати її у вологому стані, то з паренхімних клітин, що є в ній, або з калюсу, що утворюється на поверхні зрізу, часто розвиваються нові коріння; нирка при цьому продовжує рости і дає початок новій рослині. Те саме відбувається в природі, коли відламується гілка. Плеті та столони поділяються в результаті відмирання старих ділянок (міжвузлів). Так само розділяються кореневища ірису, вовчої стопи чи папороті, утворюючи нові рослини. Зазвичай бульби, наприклад бульби картоплі, продовжують жити після відмирання підземного стебла, де вони виросли; з настанням нового вегетаційного періоду вони можуть дати початок власним корінням і пагонам. У цибулинних рослин, наприклад, у гіацинтів чи тюльпанів, пагони формуються біля основи лус цибулини і можуть у свою чергу утворювати нові цибулини, які зрештою дають коріння і квітконосні стебла, тобто. стають самостійними рослинами. У деяких лілейних повітряні цибулинки утворюються в пазухах листя, а у ряду папоротей на листі виростають виводкові бруньки; Якоїсь миті вони опадають на землю і відновлюють зростання. Коріння менш здатне до утворення нових частин, ніж стебла. Бульбу жоржини для цього необхідна нирка, що утворюється біля основи стебла; проте батат може дати початок новій рослині з нирки, що утворюється кореневою шишкою. Листя теж здатне до регенерації. У деяких видів папоротей, наприклад, у кривокучника (Camptosorus), листя сильно витягнуте і має вигляд довгих волосоподібних утворень, що закінчуються меристемою. З цієї меристеми розвивається зародок із зародковими стеблами, корінням та листям; якщо кінчик листа батьківської рослини нахилиться вниз і доторкнеться до землі або моху, зачаток починає рости. Нова рослина відокремлюється від батьківського після виснаження цієї волосоподібної освіти. Листя сукулентної кімнатної рослини каланхое несуть по краях добре розвинені рослинки, які легко відпадають. Нові пагони та коріння формуються на поверхні листя бегонії. Спеціальні тільця, які називаються зародковими нирками, розвиваються на листі деяких плаунових (Lycopodium) та печіночників (Marchantia); впавши на землю, вони укорінюються та утворюють нові зрілі рослини. Багато водоростей успішно розмножуються, розчленовуючи на фрагменти під ударами хвиль.
Див. такожСИСТЕМАТИКА РОСЛИН. ЛІТЕРАТУРА Меттсон П. Регенерація - сьогодення та майбутнє. М., 1982. Гілберт С. Біологія розвитку, тт. 1-3. М., 1993-1995
Енциклопедія Кольєра. - Відкрите суспільство. 2000 .
Синоніми:Дивитись що таке "РЕГЕНЕРАЦІЯ" в інших словниках:
РЕГЕНЕРАЦІЯ- РЕГЕНЕРАЦІЯ, процес утворення нового, органу чи тканини дома віддаленого тим чи іншим чином ділянки організму. Дуже часто Р. визначається як процес відновлення втраченого, тобто утворення органу, подібного до віддаленого. Таке… … Велика медична енциклопедія
- (пізн. лат., від лат. re знову, знову, і genus, eris рід, покоління). Відродження, поновлення, відновлення того, що було зруйновано. У фігуральному значенні: зміна на краще. Словник іноземних слів, що увійшли до складу російської мови. Словник іноземних слів російської мови
РЕГЕНЕРАЦІЯ, в біології здатність організму до заміщення однієї з втрачених елементів. Термін регенерація також відноситься до форми БЕЗПЕЧНОГО РОЗМНОЖЕННЯ, при якому нова особина виникає з відокремленої частини материнського організму. Науково-технічний енциклопедичний словник
Відновлення, рекуперація; відшкодування, регенерування, відновлення, гетероморфоз, петтенкоферування, відродження, морфалаксис Словник російських синонімів. відновлення сут., кіл у синонімів: 11 відшкодування (20) … Словник синонімів
1) відновлення за допомогою певних фізико-хімічних процесів вихідного складу та властивостей відпрацьованих продуктів для повторного їх використання. У військовій справі широкого поширення набула регенерація повітря (особливо на підводних морських словниках.
Регенерація- - Повернення відпрацьованому продукту вихідних властивостей. [Термінологічний словник з бетону та залізобетону. ФГУП «НДЦ «Будівництво» НДІЗБ ім. А. А. Гвоздєва, Москва, 2007 110 стор.] Регенерація - відновлення відпрацьованих ... ... Енциклопедія термінів, визначень та пояснень будівельних матеріалів
РЕГЕНЕРАЦІЯ- (1) відновлення вихідних властивостей та складу відпрацьованих матеріалів (води, повітря, масел, гуми та ін.) для їх повторного використання. Здійснюється за допомогою певних фіз. хім. процесів у спеціальних пристроях регенераторах. Широко… … Велика політехнічна енциклопедія
- (від позднелат. regeneratio відродження відновлення), в біології відновлення організмом втрачених або пошкоджених органів і тканин, а також відновлення цілого організму з його частини. Більшою мірою властива рослинам і безхребетним.
У техніці,1) повернення продукту, що відпрацював, вихідних якостей, напр. відновлення властивостей відпрацьованої формувальної суміші в ливарному виробництві, очищення мастила, що відпрацювало, перетворення зношених гумових виробів на пластичну… Великий Енциклопедичний словник
РЕГЕНЕРАЦІЯ, регенерації, багато інших. ні, дружин. (Лат. regeneratio відновлення, повернення). 1. Нагрів газу та повітря, що надходять у піч, відпрацьованими продуктами горіння (тех.). 2. Відтворення тваринами втрачених органів (зоол.). 3. Випромінювання… … Тлумачний словник Ушакова
Регенерація(Від лат. regeneratio- Відродження) - процес відновлення біологічних структур в ході життєдіяльності організму. Регенерація підтримує будову та функції організму, його цілісність. Регенераційні процеси реалізуються на різних рівнях організації - молекулярно-генетичному, субклітинному, клітинному, тканинному, органному, організмовому. і т.д. Всі ці процеси входять в обмін речовин клітини. На субклітинному рівні відбувається відновлення структур клітини за рахунок утворення нових структурних одиниць і складання органел або поділу органел, що збереглися. Наприклад, рухомі утворення клітинної мембрани - рецептори, іонні канали та насоси - можуть переміщатися, концентруватися або розподілятися у складі мембрани. Крім цього, вони виходять з мембрани, руйнуються і замінюються новими. Так, у міобластах щохвилини деградує і замінюється новими молекулами приблизно 1 мкм2 поверхні. У фоторецепторних клітинах - паличках (рис. 8.73) є зовнішній сегмент, що складається приблизно з тисячі так званих фоторецепторних дисків - щільно покладених ділянок клітинної мембрани, в які занурені світлочутливі білки, пов'язані з зоровим пігментом. Ці диски безперервно оновлюються - деградують на зовнішньому кінці і знову з'являються на внутрішньому зі швидкістю 3-4 диски на годину. Аналогічно здійснюються процеси відновлення після пошкоджень. Вплив мітохондріальних отрут викликає втрату кріст мітохондрій. Після припинення дії отрути в клітині печінки мітохондрії відновлюють свою структуру за 2-3 сут.Клеточный рівень регенерації передбачає відновлення структури і, у деяких випадках, функцій клітини. Наприклад такого роду відноситься відновлення відростка нервової клітини нейрона. У ссавців цей процес йде зі швидкістю 1 мм на добу. Відновлення функцій клітини може здійснюватись за рахунок гіперплазії- збільшення кількості внутрішньоклітинних органел (внутрішньоклітинна регенерація). На наступному рівні - тканинному або клітинно-популяційному - відбувається заповнення клітин певного напрямку диференціювання, що втрачаються. Відбуваються перебудови не більше клітинних по-пуляций, та його результатом стає відновлення функцій тканини. Так, у людини час життя клітин кишкового епітелію - 4-5 діб, тромбоцитів - 5-7 діб, еритроцитів - 120-125 діб. Щомиті руйнується близько 1 млн еритроцитів і стільки ж утворюється в червоному кістковому мозку знову. Можливість відновлення втрачених клітин забезпечується завдяки тому, що в тканинах існує два клітинні компартменти. Один - диференційовані робочі клітини, а інший - камбіальні клітини, здатні до поділу та подальшого диференціювання. Ці останні нині називають регіональними стовбуровими клітинами (див. пп. 3.1.2, 3.2). Вони коміті-ровані, тобто. доля їх зумовлена (див. п. 8.3.1), тому вони здатні дати початок одному або декільком певним клітинним типам. Їх подальше диференціювання визначається сигналами, що надходять ззовні: від оточення (міжклітинними взаємодіями) та дистантними (наприклад, гормонами), залежно від яких у клітинах вибірково активуються конкретні гени. Так, в епітелії тонкої кишки камбіальні клітини перебувають у придонних зонах крипт (рис. 8.74). При певних впливах вони здатні дати початок клітинам «каємчастого» епітелію, що всмоктує, і деяким одноклітинним залозам. Органний рівень регенерації передбачає відновлення функції або структури органу. У цьому рівні спостерігаються як перетворення клітинних популяцій, а й морфогенетичні процеси. У цьому реалізуються самі механізми, як і для формуванні органів в эмбриогенезе. Та- Мал. 8.73.Схематичне зображення фоторецептора сітківки - палички: 1 - синаптичне тільце, що примикає до нейрального шару сітківки, 2 - ядро, 3 - апарат Гольджі, 4 - внутрішній сегмент з мітохондріями, 5 - сполучна війка, 6 - зовнішній сегмент яка регенерація може здійснюватися шляхомепіморфозу, морфолаксису, регенераційну гіпертрофію.Ціспособи та механізми регенерації обговорюються далі.На організмовому рівні можливе окремих випадках відтворення цілісного організму з однієї чи групи клітин. Розрізняють два види регенерації:фізіологічнуірепаративну.Фізіологічна (гомеостатична) регенераціяє процес відновлення структур, які зношуються в процесі нормальної життєдіяльності. Завдяки їй підтримується структурний гомеостаз та забезпечується можливість постійного виконання органами їх функцій. З загальнобіологічної погляду фізіологічна регенерація, як і обмін речовин, є проявом такого найважливішого якості життя, як самооновлення. Самооновлення забезпечує існування організму у часі та просторі. У його основі лежить біогенна міграція атомів. На внутрішньоклітинному рівні значення фізіологічної регенерації особливо велике для про «вічних» тканин, які втратили здатність до регенерації шляхом поділу клітин. Насамперед це стосується нервової тканини, сітківки ока. На клітинному та тканинному рівнях здійснюється фізіологічна регенерація у «лабільних» тканинах, де Мал. 8.74.Локалізація регіональних стовбурових клітин в епітелії тонкої кишки: 1 - клітини, що не діляться; 2 — стовбурові клітини, що діляться; 3 — клітини, що швидко діляться; 4 - диференційовані клітини, що не діляться; 5 - напрямок переміщення клітин; 6 — клітини, злущені з поверхні кишкової ворсини, інтенсивність клітинного оновлення дуже велика, і в «зростаючих» тканинах, клітини яких оновлюються значно повільніше. До першої групи належать, наприклад, рогівка ока, епітелій слизової оболонки кишечника, клітини периферичної крові, епідерміс шкіри та його похідні – волосся та нігті. Клітини таких органів, як печінка, нирка, наднирковий залози становлять другу із зазначених груп. Про інтенсивність проліферації судять за кількістю мітозів, що припадають на 1000 підрахованих клітин. Якщо врахувати, що сам мітоз в середньому триває близько 1 години, а весь мітотичний цикл у соматичних клітинах в середньому протікає 22-24 год, стає ясно, що для визначення інтенсивності оновлення клітинного складу тканин необхідно підрахувати число мітозів протягом однієї або декількох діб. Виявилося, що кількість клітин, що діляться, не однакова в різні години доби. Так було відкрито добовий ритм клітинних поділів, приклад якого зображено на рис. 8.75.Добовий ритм числа мітозів виявлено у нормальних, а й у пухлинних тканинах. Він відбиває більш загальну закономірність, Мал. 8.75.Добові зміни мітотичного індексу (МІ) в епітелії стравоходу (1) і рогівки (2) мишей. Мітотичний індекс виражений у проміле (0/00), що відображає кількість мітозів у тисячі підрахованих клітин зокрема, ритмічність всіх функцій організму. Одна з сучасних галузей біології.хронобіологія— вивчає зокрема механізми регуляції добових ритмів мітотичної активності, що має дуже велике значення для медицини. Існування добової періодичності числа мітозів вказує на регульованість фізіологічної регенерації організмом. Крім добових, існують місячні та річні цикли оновлення тканин та органів. Фізіологічна регенерація властива організмам всіх видів, але особливо інтенсивно вона протікає у теплокровних хребетних, оскільки вони взагалі дуже висока інтенсивність функціонування всіх органів проти іншими тваринами. Репаративна регенерація(Від лат.reparatio — відновлення) — відновлення біологічних структур після травм та дії інших факторів, що ушкоджують. До таких факторів можуть бути віднесені отруйні речовини, хвороботворні агенти, високі та низькі температури (опіки та обмороження), променеві дії, голодування тощо. Здатність до регенерації не має однозначної залежності від рівня організації, хоча давно вже було помічено, що більш низько організовані тварини мають кращу здатність до регенерації зовнішніх органів. Це підтверджується дивовижними прикладами регенерації гідри, планарій, кільчастих черв'яків, членистоногих, голкошкірих, нижчих хордових, наприклад, асцидій. З хребетних найкращою регенераційною здатністю мають хвостаті земноводні. Відомо, що різні види того самого класу можуть сильно відрізнятися за здатністю до регенерації. Крім того, при вивченні здатності до регенерації внутрішніх органів виявилося, що вона значно вища у теплокровних тварин, наприклад, у ссавців, порівняно із земноводними. Регенерація у ссавців відрізняється своєрідністю. Для регенерації деяких зовнішніх органів необхідні спеціальні умови. Мова, вухо, наприклад, не регенерують при крайовому ушкодженні (фактично йдеться про ампутацію крайової частини структури). Якщо ж нанести наскрізний дефект через усю товщу органа, відновлення йде добре. Регенерація внутрішніх органів може дуже активно. З невеликого фрагмента яєчника поновлюється цілий орган. Є припущення, що неможливість регенерації кінцівок та інших зовнішніх органів у ссавців має пристосувальний характер і зумовлена відбором, оскільки при активному способі життя морфогенетичні процеси, що вимагають складної регуляції, ускладнювали б існування. Ряд дослідників вважає, що організми спочатку мали два методи лікування від ран — вплив імунної системи та регенерацію. Але в ході еволюції вони стали несумісними один з одним. Хоча регенерація може бути кращим вибором, для нас важливіші Т-клітини імунної системи — основна зброя проти пухлин. Регенерація кінцівки стає безглуздою, якщо водночас у організмі бурхливо розвиваються ракові клітини. Виходить, що імунна система, захищаючи нас від інфекцій та раку, одночасно пригнічує наші здібності до відновлення. Обсяг репаративної регенерації може бути дуже різним. Крайній варіант – відновлення цілого організму з окремої малої його частини, фактично з групи соматичних клітин. Серед тварин таке відновлення можливе у губок та кишковопорожнинних. Регенерацію гідри можна здійснити із групи клітин, отриманих при продавлюванні її через сито. Серед рослин можливий розвиток цілої нової рослини навіть із однієї соматичної клітини, як це отримано на прикладі моркви та тютюну. Такий вид відновлювальних процесів супроводжується виникненням нової морфогенетичної осі організму та названий Б.П. Токіним «соматичним ембріогенезом», оскільки багато в чому нагадує ембріональний розвиток. Як подібний варіант регенерації може розглядатися клонування в експерименті цілого організму з однієї соматичної клітини у ссавців. Наступний за обсягом варіант — відновлення великих ділянок організму, що складаються з комплексу органів. Прикладом є регенерація у гідри, війного хробака (планарії), морської зірки (рис. 8.76). При видаленні частини тварини з фрагмента, що залишився, навіть дуже невеликого, можливе відновлення повноцінного організму. Наприклад, відновлення морської зірки з променя, що зберігся. кісток та інших внутрішніх органів - найменш об'ємний процес, але не менш важливий для відновлення структурно-функціональної цілісності організму. Існує кілька способів репаративної регенерації. До них відносять епіморфоз, морфалаксис, регенераційну гіпертрофію, компенсаторну гіпертрофію, загоєння епітеліальних ран, тканинну регенерацію. Мал. 8.76.Регенерація комплексу органів деяких видів безхребетних тварин: а — гідра; б - плоский черв'як; в - морська зірка; г - відновлення морської зірки з променя Епіморфозє найбільш очевидним способом регенерації, що полягає у відростанні нового органу від ампутаційної поверхні. Ілюстрацією може бути регенерація кришталика чи кінцівки у хвостатих амфібій (рис. 8.77). Розглянемо детальніше процес регенерації з прикладу эпиморфоза кінцівки тритона. У процесі відновлення виділяють регресивну та прогресивну фази регенерації. Регресивна фаза починається із загоєння рани, під час якого відбуваються такі основні події: зупинка Мал. 8.77.Регенерація кришталика (1) з дорзальної райдужної оболонки (2) у тритонакровотечі, скорочення м'яких тканин кукси кінцівки, освіта над раневою поверхнею згустку фібрину і міграція епідермісу, що покриває ампутаційну поверхню. Потім починається руйнування тканин безпосередньо проксимальніше місця ам. Одночасно в зруйновані м'які тканини проникають клітини, що у запальному процесі, спостерігається фагоцитоз і місцевий набряк. Після цього в області під рановим епідермісом починається дедиференціювання спеціалізованих клітин: м'язових, кісткових, хрящових тощо. Клітини набувають рис мезенхімних, утворюють скупчення і формують регенераційну бластему(Рис. 8.78). У цей час рановий епідерміс швидко потовщується і утворює апікальну ектодермальну шапочку.На цьому етапі в регенераційну бластему та ектодермальну шапочку вростають судини та нервові волокна. Далі починається прогресивна фаза, для якої найбільш характерні процеси росту та морфогенезу. Довжина та маса регенераційної бластеми швидко збільшуються. Вона набуває конічної форми. Мезенхімні клітини бластеми дедиференціюються, даючи початок всім спеціалізованим клітинним типам, які необхідні для формування структур кінцівки. Здійснюється зростання кінцівки та її морфогенез (формоутворення). Коли форма кінцівки в загальних рисах вже склалася, регенерат все ще менший за нормальну кінцівку. Чим більша тварина, тим більша ця різниця в розмірах. Для завершення морфогенезу потрібен час, після якого регенерат досягає розмірів нормальної кінцівки. Деякі стадії відновлення передньої кінцівки у тритону після ампутації на рівні плеча показані на рис. 8.79. Мал. 8.78.Регенерація кінцівки тритону: а — нормальна кінцівка, б — ампутація; в - формування апікальної шапочки та бластеми; г - редиференціювання клітин; д - знову сформована кінцівка. 1 - бластема; 2 - апікальна ектодермальна шапочка; 3 – редиференціювання клітин бластеми (пояснення в тексті) У молодих личинок аксолотлів кінцівка може регенерувати за 3 тижні, у дорослих тритонів та аксолотлів – за 1-2 міс, а у наземних амбістів для цього потрібно близько 1 року. Морфалаксис- Регенерація шляхом перебудови ділянки, що регенерує. Прикладом служить регенерація гідри з кільця, вирізаного з середини тіла, або відновлення планарії з однієї десятої або двадцятої її частини. На раневой поверхні у разі немає значних формообразовательных процесів. Відрізаний шматочок стискається, клітини всередині нього перебудовуються, і виникає ціла особина зменшених розмірів, яка зростає. Цей спосіб регенерації вперше описав Т. Морган у 1900 р. Відповідно до його опису, морфалаксис здійснюється без мітозів. Нерідко має місце поєднання епіморфного зростання на місці ампутації з реорганізацією шляхом морфалаксису в прилеглих частинах тіла. Регенераційна гіпертрофія (ендоморфоз)відноситься до внутрішніх органів. Цей спосіб регенерації полягає у збільшенні розмірів залишку органу без відновлення вихідної форми. Ілюстрацією служить регенерація печінки хребетних, у тому числі ссавців. При крайовому пораненні печінки віддалена частина органу ніколи не відновлюється. Ранева поверхня гоїться. У той же час вну- Мал. 8.79.Регенерація передньої кінцівки тритону в експерименті Мал. 8.80.Вплив віку збільшення числа клубочків нефронів після видалення однієї нирки у щурів невдовзі після народження: 1 — крива приросту числа клубочків у нормальному постнатальном розвитку лише у нирці; 2 — криві збільшення числа новостворених клубочків після видалення нирки на різних термінах онтогенезатри частини, що залишилася, посилюється розмноження клітин (гіперплазія) і навіть після видалення 2/3 печінки відновлюються вихідні маса і об'єм, але не форма. Внутрішня структура печінки виявляється нормальною, часточки мають типову їм величину. Функція печінки також повертається до норми. Компенсаторна (вікарна) гіпертрофіяполягає у змінах в одному з органів при порушенні в іншому, що відноситься до тієї самої системи органів. Приклад - гіпертрофія в одній із нирок при видаленні іншої або збільшення лімфатичних вузлів при видаленні селезінки. Зміни здатності до такого типу регенерації в залежності від віку показано на рис. 8.80. Останні два способи відрізняються місцем регенерації, але механізми їх однакові: гіперплазія та гіпертрофія (рис. 8.81)1. 1 Гіпертрофія(грец. hyper-+ trophe— їжа, харчування)- Збільшення об'єму і маси органу тіла або окремої його частини. Гіперплазія (грец. hyper-+ plasis- Освіта, формування) - збільшення числа структурних елементів тканин шляхом їх надлишкового новоутворення. Це не тільки розмноження клітин, а й збільшення цитоплазматичних ультраструктур (змінюються в першу чергу мітохондрії, міофіламенти, ендоплазматичний ретикулум, рибосоми). Мал. 8.81.Схема, що ілюструє механізми гіпертрофії та гіперплазії: а - норма; б - гіперплазія; в - гіпертрофія; г - комбінована зміна Епітелізаціяпри загоєнні ран з порушеним епітеліальним покривом йде приблизно однаково, незалежно від того, далі відбуватиметься регенерація органу шляхом епіморфозу чи ні. Епідермальне загоєння рани у ссавців у тому випадку, коли ранова поверхня висихає з утворенням кірки, проходить наступним чином (рис. 8.82). Епітелій на краю рани потовщується внаслідок збільшення об'єму клітин та розширення міжклітинних просторів. Потік фібрину відіграє роль субстрату для міграції епідермісу в глиб рани. У мігруючих епітеліальних клітинах немає мітозів, одна- Мал. 8.82.Схема деяких подій, що відбуваються при епітелізації шкірної рани у ссавців: а - початок вростання епідермісу під некротичну тканину; б - зрощення епідермісу та відділення струпа; 1 - сполучна тканина; 2 - епідерміс; 3 - струп; 4 - некротична тканина вони мають фагоцитарну активність. Клітини з протилежних країв входять у контакт. Потім настає кератинізація ранового епідермісу та відділення кірки, що покриває рану. До моменту зустрічі епідермісу протилежних країв у клітинах, розташованих безпосередньо навколо краю рани, спостерігається спалах мітозів, який потім поступово згасає. Відновлення окремих мезодермальних тканин, таких як м'язова та скелетна, називають тканинної регенерації.Для регенерації м'яза важливе збереження хоча б невеликих її кукс на обох кінцях, а для регенерації кістки необхідна окістя. Таким чином, існує безліч різних способів або типів морфогенетичних явищ при відновленні втрачених і пошкоджених частин організму. Відмінності між ними не завжди очевидні, і потрібне глибше розуміння цих процесів. При регенерації не завжди утворюється точна копія віддаленої структури. В разі типовоїрегенерації відновлюється втрачена частина правильної структури (гомоморфоз),чого не відбувається за атиповійрегенерації. Прикладом останньої є поява іншої структури на місці втраченої. гетероморфоз.Вона може виявлятися у вигляді гомеозноїрегенерації, що полягає у появі антени або кінцівки на місці ока у членистоногих. Ще один варіант - гіпоморфоз,регенерація із частковим заміщенням ампутованої структури. Наприклад, у ящірки виникає шилоподібна структура замість кінцівки (рис. 8.83). До атипової регенерації можуть бути віднесені випадки зміни полярностіструктури. Так, із короткого фрагмента планарії можна стабільно отримувати біполярну планарію. Зустрічається утворення додаткових структур або надмірна регенерація. Після надрізу кукси при ампутації головного відділу планарії виникає регенерація двох голів або більше (рис. 8.84). Вивчення регенерації стосується не лише зовнішніх проявів. Існує ціла низка аспектів, які мають проблемний і теоретичний характер. До них відносяться питання регуляції та умов, в яких протікають відновлювальні процеси, питання походження клітин, що беруть участь у регенерації, здатності до регенерації у різних груп тварин та особливостей відновлювальних процесів у ссавців. Встановлено, що при регенерації відбуваються такі процеси, як детермінація, диференціювання та диференціація, зростання, морфоге- Мал. 8.83.Приклади атипової регенерації: а – нормальна голова раку; б - формування антени замість ока; в - утворення шиловидної структури замість кінцівки у саламандри. 1 - око; 2 - антена; 3 - місце ампутації; 4 - нервовий ганглій Мал. 8.84.Приклади атипової регенерації: а – біполярна планарію; б — багатоголова планарія, отримана після ампутації голови та нанесення насічок на культюнез, подібні до процесів, що мають місце в ембріональному розвитку. Дані, отримані до теперішнього часу, вказують на те, що відновлення втрачених структур, по суті, здійснюється на основі тієї ж самої програми розвитку,яка керує формуванням їх у ембріона, та на основі клітинних та системних механізмів розвитку. Проте за регенерації процеси розвитку йдуть вже вдруге, тобто. у сформованому організмі, тому відновлення структур має низку відмінностей та специфічних рис. Безперечно, що в ході регенерації велике значення належить системним механізмам — міжклітинним та міжзачатковим взаємодіям, нервовій та гуморальній регуляції. Так, при епіморфозі кінцівки тритону сформований під час епітелізації епідерміс стимулює лізис мезодермальних тканин, що підлягають. У його відсутність або утворенні шраму регенерації немає. Клітини під сформованим епідермісом дедиференціюються та формують бластему. На цьому етапі спостерігаються реципрокні індуктивні впливи між епідермісом, який формує апікальну ектодермальну шапочку, та мезодер-мальної бластеми. У ході ембріонального розвитку при формуванні кінцівки здійснювалися подібні взаємодії між мезодермальною ниркою кінцівки та апікальним ектодермальним гребенем. У ході дедиференціювання у клітинах пригнічується активність типо-специфічних генів, що визначають спеціалізацію клітини, наприклад генів MRFіMif5у м'язових волокнах. Потім активуються гени, необхідні проліферації клітин. Один з нихmsx1.На цій стадії нервові відростки, що вростають в бластему, і епідерміс продукують трофічні і ростові фактори, необхідні для проліферації і виживання клітин бластеми. Серед них фактор зростання фібробластів FGF-10.Цей фактор необхідний для проліферації самого епідермісу. Бластема, своєю чергою, синтезує у відповідь нейротрофічні чинники, що стимулюють вростання нервів. Нерви потрібні на формування апікальної ектодермальної шапочки. Крім цього бластему, так само як і апікальна епідермальна шапочка, продукує FGF-8,що стимулює вростання капілярів.Слід зазначити спостерігаються на цій стадії відмінності між регенерацією та ембріональним розвитком. Для реалізації регенерації потрібна іннервація. Без неї може проходити дедиференціювання клітин, але подальший розвиток відсутній. У період ембріонального морфогенезу кінцівки (під час клітинних диференціювань) нерви ще сформовані. Крім іннервації на ранній стадії регенерації потрібна дія ферментів металопротеїназ. Вони руйнують компоненти матриксу, що дозволяє клітинам розділитися (дисоціювати) і активно проліферувати. Клітини, що контактують між собою, не можуть продовжувати регенерацію і відповідати на дію ростових факторів. Таким чином, у ході регенерації спостерігаються всі варіанти міжклітинних взаємодій: шляхом виділення паракринних факторів, що дифундують від однієї клітини до іншої, взаємодії через матрикс та при безпосередньому контакті клітинних поверхонь. У стадії дедиференціювання в клітинах кукси експресуються гомеозисні гениHoxD8іHoxDlO,а з початком диференціювання - гениHoxD9іHoxD13.Як було показано в п. 8.3.4, ці гени активно транскрибуються і в ембріональному морфогенезі кінцівки.Важливо відзначити, що в ході регенерації втрачається диференціювання клітин, а їх детермінація зберігається. Вже на стадії недиференційованої бластеми закладаються основні риси кінцівки, що регенерує. При цьому не потрібна активація генів, які забезпечують специфікацію кінцівки. (Tbx-5для передньої таTbx-4для задньої). Кінцівка формується залежно від локалізації бластеми. Її розвиток відбувається так само, як і в ембріогенезі: спочатку проксимальні відділи, а потім дистальні. Проксимально-дистальний градієнт, від якого залежить, які частини зростаючого зачатка стануть плечем, які передпліччям, а які пензлем, задається градієнтом білка. Prod 1.Він локалізований на поверхні клітин бластеми і його концентрація вище в основі кінцівки. Цей білок відіграє роль рецептора, а сигнальною молекулою (лігандом) для нього є білок nAG.Він синтезується шванновськими клітинами, що оточують регенеруючий нерв. За відсутності цього білка, який через ліганд-рецепторну взаємодію запускає активацію необхідного для розвитку каскаду генів, регенерації не відбувається. Це пояснює феномен відсутності відновлення кінцівки при перерізанні нерва, а також при вростанні в бластему недостатньої кількості нервових волокон. Цікаво, що й нерв кінцівки тритона відвести під шкіру підстави кінцівки, то утворюється додаткова кінцівка. Якщо його відвести до основи хвоста – стимулюється утворення додаткового хвоста. Відведення нерва на бічну ділянку ніяких додаткових структур не викликає. Все це призвело до створення концепції регенераційних полів. Мал. 8.85.Експеримент з поворотом бластеми кінцівки (пояснення в тексті) Аналогічно процесу ембріогенезу формується і передньо-задня вісь в полі кінцівки, що розвивається. У зачатку, що формується, з'являється зона поляризуючої активності, що визначає асиметрію кінцівки. Повернувши кінець кукси кінцівки на 180°, можна отримати кінцівку із дзеркальним подвоєнням пальців (рис. 8.85). Таким чином, справедливо твердження, що формування кінцівки відбувається в полі органу, а бластема є системою, що саморегулюється. Поряд із вищесказаним, доказом цього є результати, отримані в серії експериментів з пересадки бластеми передньої кінцівки на бластему середини стегна (рис. 8.86). При пересадці в регенераційне поле іншої кінцівки трансплантат розташовується відповідно до отриманої позиційної інформації (градієнти речовин): бластема плеча зміщується до середини стегна, передпліччя - до гомілки, зап'ястя - до лапки. Розвиток трансплантованої бластеми у відповідну частину передньої кінцівки відбувається відповідно до її детермінації, яка визначається рівнем ампутації. Це цілком зрозуміло тим, що регенерація здійснюється у вже сформованому організмі, де основними регулюючими механізмами є останні. Серед гуморальних впливів слід зупинитись на дії гормонів. Альдостерон, гормони щитовидної залози та гіпофіза надають стимулюючий вплив на відновлення втрачених Мал. 8.86.Досліди з пересадки бластеми передньої кінцівки в полі задньої (пояснення в тексті) структур. Подібну дію мають і метаболіти, які виділяються пошкодженою тканиною і транспортуються плазмою крові або передаються через міжклітинну рідину. Саме тому додаткове ушкодження у деяких випадках прискорює процес регенерації. Крім перерахованого на регенерацію впливають інші чинники, серед яких температура, при якій відбувається відновлення, вік тварини, функціонування органу, що стимулює регенерацію, і в певних ситуаціях зміна електричного заряду в регенераті. Встановлено, що в кінцівці амфібій після ампутації та в процесі регенерації відбуваються реальні зміни електричної активності. При проведенні електричного струму через ампутовану кінцівку у дорослих жаб шпорця спостерігається посилення регенерації передніх кінцівок. У регенератах збільшується кількість нервової тканини, з чого робиться висновок, що електричний струм стимулює вростання нервів у краї кінцівок, які в нормі не регенерують. Спроби стимулювати подібним чином відновлення кінцівок у ссавців виявилися безуспішними. Під дією електричного струму або при поєднанні дії електричного струму з фактором росту нервів вдавалося отримати у щура тільки розростання скелетної тканини у вигляді хрящових і кісткових мозолів, які не були схожі на нормальні елементи скелета кінцівок. Одне з найбільш інтригуючих теоретично регенерації — питання її клітинних джерелах. Звідки беруться чи як виникають недиференційовані клітини бластеми, морфологічно подібні до мезен-хімних? Нині говорять про три можливіджерела регенерації.Перший - цедедиференційовані клітини,другий -регіональні стовбурові клітиниі третій -стовбурові клітини з інших структур,які мігрували до місця регенерації.Більшість дослідників визнають дедиференціювання та метаплазію при регенерації кришталика у амфібій. Теоретичне значення цієї проблеми полягає в допущенні можливості або неможливості змін клітиною її програми настільки, що вона повертається в стан, коли знову здатна ділитися і репрограмувати свій синтетичний апарат. Наявність регіональних стовбурових клітин встановлено на даний час у багатьох тканинах: у м'язах, кістці, епідермісі шкіри, печінці, сітківці та інших. Такі клітини виявлені навіть у нервовій тканині – у певних зонах головного мозку. У багатьох випадках вважають, що джерелом, з якого утворюються диференційовані клітини в ході регенерації, є вони (регенеративна медицина, регенеративна ветеринарія). Передбачається, що зі збільшенням віку особини чисельність популяцій регіональних стовбурових клітин скорочується. Якщо ж в органі не вистачає своїх регіональних стовбурових клітин, то в нього можуть мігрувати клітини з інших та дати початок потрібній тканині. Нещодавно показано, що стовбурові клітини ізольовані з однієї дорослої тканини можуть дати початок зрілим клітинам інших клітинних ліній, незалежно від призначення класичного зародкового шару. Так, ендотелій великих магістральних артерій немає власних запасів стовбурових клітин. Його оновлення відбувається за рахунок стовбурових клітин кісткового мозку, що надходять у кровотік. Проте порівняльна неефективність таких перетворень in vivo(в організмі), навіть за наявності пошкодження тканини, ставить питання про те, чи має цей механізм фізіологічне значення. .Якщо вдасться вирішити питання трансформації клітинних ліній, то цілком можливим стане використання цих технологій в репаративної медицині для лікування широкого кола хвороб. Однак, незважаючи на досягнення біології останніх років, у проблемі регенерації ще залишається дуже багато невирішених питань.
Регенерація (в патології) - це відновлення цілості тканин, порушеної будь-яким хворобливим процесом або зовнішнім впливом, що травмує. Відновлення відбувається за рахунок сусідніх клітин, заповнення молодими клітинами дефекту та подальшого перетворення їх на зрілу тканину. Така форма носить назву репаративної (що відшкодовує) регенерації. При цьому можливі два варіанти регенерації: 1) спад відшкодовується тканиною того ж виду, що і загибла (повна регенерація); 2) спад заміщається молодою сполучною (грануляційною) тканиною, що перетворюється на рубцеву (неповна регенерація), що є не регенерацією у власному розумінні, а загоєнням тканинного дефекту.
Регенерація передує звільнення цієї ділянки від загиблих клітин шляхом ферментативного їх розплавлення та всмоктування в лімфу або кров або шляхом (див.). Продукти розплавлення є одним із стимуляторів розмноження сусідніх клітин. У багатьох органах та системах існують ділянки, клітини яких є джерелом розмноження клітин при регенерації. Наприклад, у кістковій системі таким джерелом є окістя, клітини якої, розмножуючись, утворюють спочатку остеоїдну тканину, що надалі перетворюється на кістку; у слизових оболонках – клітини глибоколежачих залоз (крипти). Регенерація клітин крові відбувається в кістковому мозку та поза ним у системі та її похідних (лімфатичних вузлах, селезінці).
Здатність до регенерації мають далеко не всі тканини і не однаковою мірою. Так, м'язові клітини серця не здатні до розмноження, що завершується утворенням зрілих м'язових волокон, тому будь-який дефект м'язів міокарда заміщується рубцем (зокрема після інфаркту). При загибелі тканини мозку (після крововиливу, артеріосклеротичного розм'якшення) дефект не замінюється нервовою тканиною, а утворюється кіота.
Іноді виникає при регенерації тканина за будовою відрізняється від вихідної (атипова регенерація) або об'єм її перевищує обсяг загиблої тканини (гіперрегенерація). Такий перебіг регенераційного процесу може призвести до виникнення пухлинного росту.
Регенерація (лат. regenerate – відродження, відновлення) – відновлення анатомічної цілості органа або тканини після загибелі структурних елементів.
У фізіологічних умовах процеси регенерації відбуваються безперервно з різною інтенсивністю в різних органах і тканинах відповідно до інтенсивності відживання клітинних елементів даного органу або тканини і заміщення їх новоствореними. Безперервно заміщаються формені елементи крові, клітини покривного епітелію шкіри, слизових оболонок шлунково-кишкового тракту, дихальних шляхів. Циклічні процеси в жіночій статевій сфері призводять до ритмічного відторгнення та оновлення ендометрію шляхом його регенерації.
Усі ці процеси є фізіологічним прототипом патологічної регенерації (її називають ще репаративною). Особливості розвитку, перебігу та результату репаративної регенерації визначаються розмірами загибелі тканини та характером патогенних впливів. Остання обставина особливо треба мати на увазі, оскільки умови та причини загибелі тканини мають суттєве значення для регенераційного процесу та його наслідків. Так, наприклад, особливий характер мають рубці після опіків шкіри, що відрізняються від рубців іншого походження; сифілітичні рубці грубі, призводять до глибоких втягувань та спотворення органу і т. д. На відміну від фізіологічної регенерації, репаративна регенерація охоплює широке коло процесів, що ведуть до відшкодування дефекту, викликаного втратою тканини внаслідок її ушкодження. Розрізняють повну репаративну регенерацію - реституцію (заміщення дефекту тканиною того ж виду і тієї ж структури, що і загибла) і неповну репаративну регенерацію (заповнення дефекту тканиною, що володіє більшими пластичними властивостями, ніж загибла, тобто звичайною грануляційною тканиною подальшим перетворенням її на рубцеву). Таким чином, у патології під регенерацією часто мають на увазі загоєння.
З поняттям регенерації пов'язане також поняття про організацію, оскільки в основі обох процесів лежать загальні закономірності новоутворення тканин і поняття субституції, тобто витіснення та заміщення новоствореної тканини передіснувала тканини (наприклад, субституція тромбу фіброзною тканиною).
Ступінь повноти регенерації визначається двома основними факторами: 1) регенераційна потенція цієї тканини; 2) обсяг дефекту та одно- або різнорідність видової приналежності загиблих тканин.
Перший чинник нерідко пов'язують із ступенем диференціювання цієї тканини. Однак саме поняття диференціювання та зміст цього поняття є досить відносними, і порівняння тканин за цією ознакою із встановленням кількісної градації диференційованості у функціональному та морфологічному стосунках неможливе. Поряд з тканинами, що мають високу регенераційну потенцію (наприклад, тканина печінки, слизові оболонки шлунково-кишкового тракту, органи кровотворення та ін.), існують органи з нікчемною потенцією до регенерації, в яких регенерація ніколи не завершується повним відновленням втраченої тканини (наприклад, , ЦНС). Надзвичайно високою пластичністю мають сполучну тканину, елементи стінки найдрібніших кровоносних і лімфатичних судин, периферичні нерви, ретикулярну тканину та її похідні. Тому пластичне роздратування, яким є травма у сенсі цього терміну (тобто. всі форми її), передусім і найповніше стимулює зростання цих тканин.
Обсяг загиблої тканини має важливе значення для повноти регенерації, і емпірично більш менш відомі кількісні межі втрати тканини для кожного органу, що визначають ступінь відновлення. Вважають, що для повноти регенерації значення має не тільки обсяг як суто кількісна категорія, а й комплексне розмаїття загиблих тканин (це особливо стосується загибелі тканин, спричиненої токсикоінфекційними впливами). Для пояснення цього факту слід, мабуть, звернутися до загальних закономірностей стимуляції пластичних процесів в умовах патології: стимуляторами є продукти тканинної загибелі (гіпотетичні «некрогормони», «мітогенетичні промені», «трефони» тощо). Одні є специфічними стимуляторами для клітин певного виду, інші - неспецифічними, стимулюючими найбільш пластичні тканини. До неспецифічних стимуляторів відносять продукти розпаду та життєдіяльності лейкоцитів. Їх наявність при реактивному запаленні, що розвивається завжди при загибелі не тільки паренхіматозних елементів, а й судинної строми, сприяє розмноженню найбільш пластичних елементів - сполучної тканини, тобто розвитку в кінцевому рахунку рубця.
Існує загальна схема послідовності процесів регенерації незалежно від галузі, де вона відбувається. В умовах патології процеси регенерації у вузькому значенні цього слова та процеси загоєння мають різний характер. Ця різниця визначається характером загибелі тканини та вибірковою спрямованістю дії патогенного фактора. Чисті форми регенерації, т. е. відновлення тканини, ідентичної втраченої, спостерігаються у випадках, коли під впливом патогенного впливу гинуть лише специфічні паренхіматозні елементи органу за умови їх високої регенеруючої потенції. Прикладом є регенерація епітелію канальців нирки, вибірково пошкодженого токсичним впливом; регенерація епітелію слизових оболонок при десквамації його; регенерація альвеолоцитів легені при десквамативному катарі; регенерація епітелію шкіри; регенерація ендотелію кровоносних судин і ендокарда та ін. У цих випадках джерелом регенерації є клітинні елементи, розмноження, дозрівання і диференціація яких призводить до повного заміщення втрачених паренхіматозних елементів. При загибелі складних структурних комплексів відновлення втраченої тканини йде з спеціальних ділянок органу, є своєрідними центрами регенерації. У слизовій оболонці кишечника, в ендометрії такими центрами є залізисті крипти. Їх клітини, що розмножуються, покривають дефект спочатку одним шаром недиференційованих клітин, з яких потім диференціюються залози і відновлюється структура слизової. У кістковій системі таким центром регенерації є окістя, у покривному плоскому епітелії – мальпігієвий шар, у системі крові – кістковий мозок та екстрамедулярні похідні ретикулярної тканини.
Загальним законом регенерації є закон розвитку, за яким у процесі новоутворення виникають молоді недиференційовані клітинні похідні, надалі проходять етапи морфологічної і функціональної диференціювання до формування зрілої тканини.
Загибель ділянок органу, які з комплексу різних тканин, викликає реактивне запалення (див.) по периферії. Це пристосувальний акт, оскільки запальна реакція супроводжується гіперемією та підвищенням тканинного метаболізму, що сприяє зростанню новоутворюваних клітин. Крім того, клітинні елементи запалення групи гістофагоцитів є пластичним матеріалом для новоутворення сполучної тканини.
У патології нерідко анатомічне загоєння досягається за допомогою грануляційної тканини (див.) – етапу новоутворення фіброзного рубця. Грануляційна тканина розвивається майже при будь-якій репаративної регенерації, але ступінь її розвитку та кінцеві наслідки варіюють у дуже широких межах. Іноді це важко розрізнити при мікроскопічному дослідженні ніжні ділянки фіброзної тканини, іноді грубі щільні тяжі гіалінізованої брадитрофної рубцевої тканини, що нерідко піддається кальцинозу (див.) і осифікації.
Крім регенераційної потенції даної тканини, характеру її ураження, обсягу його, важливе значення у регенераційному процесі мають загальні фактори. До них відносять вік суб'єкта, характер та особливості харчування, загальну реактивність організму. При порушеннях іннервації, авітамінозах звичайний перебіг репаративної регенерації перекручується, що найчастіше виявляється у уповільненні процесу регенерації, млявості клітинних реакцій. Існує також поняття про фібропластичний діатез як про конституційну особливість організму реагувати на різні патогенні роздратування підвищеним утворенням фіброзної тканини, що проявляється формуванням келоїду (див.), спайковою хворобою. У клінічній практиці важливо враховувати загальні фактори для створення оптимальних умов повноти регенераційного процесу та загоєння.
Регенерація є одним із найважливіших пристосувальних процесів, що забезпечують відновлення здоров'я та продовження життя за надзвичайних обставин, що створюються хворобою. Однак, як і будь-який пристосувальний процес, регенерація на відомому етапі та при деяких шляхах розвитку може втрачати пристосувальне значення та сама створювати нові форми патології. Рубці, що знеображують, деформують орган, різко порушують його функцію (наприклад, рубцеве перетворення клапанів серця в результаті ендокардиту), створюють нерідко важку хронічну патологію, що вимагає спеціальних лікувальних заходів. Іноді новостворена тканина кількісно перевищує обсяг загиблої (суперрегенерація). Крім того, у кожному регенераті є елементи атипізму, різка вираженість яких є етапом розвитку пухлини (див.). Регенерація окремих органів і тканин - див. у відповідних статтях про органи та тканини.
Загальні відомості
Регенерація(Від лат. regeneratio -відродження) - відновлення (відшкодування) структурних елементів тканини замість загиблих. У біологічному сенсі регенерація є пристосувальний процес, вироблений у ході еволюції та властивий всьому живому. У життєдіяльності організму кожне функціональне відправлення потребує витрат матеріального субстрату та його відновлення. Отже, при регенерації відбувається самовідтворення живої матерії,причому це самовідтворення живого відбиває принцип ауторегуляціїі автоматизації життєвих відправлень(Давидовський І.В., 1969).
Регенераторне відновлення структури може відбуватися на різних рівнях – молекулярному, субклітинному, клітинному, тканинному та органному, проте завжди йдеться про відшкодування структури, яка здатна виконувати спеціалізовану функцію. Регенерація – це відновлення як структури, і функції.Значення регенераторного процесу – у матеріальному забезпеченні гомеостазу.
Відновлення структури та функції може здійснюватися за допомогою клітинних або внутрішньоклітинних гіперпластичних процесів. На цій підставі розрізняють клітинну та внутрішньоклітинну форми регенерації (Саркісов Д.С., 1977). Для клітинної формирегенерації характерно розмноження клітин мітотичним і амітотичним шляхом, внутрішньоклітинної форми,яка може бути органоїдною та внутрішньоорганоїдною, - збільшення числа (гіперплазія) та розмірів (гіпертрофія) ультраструктур (ядра, ядерців, мітохондрій, рибосом, пластинчастого комплексу і т.д.) та їх компонентів (див. рис. 5, 11, 15) . Внутрішньоклітинна формарегенерації є універсальною, оскільки вона властива всім органам та тканинам. Однак структурно-функціональна спеціалізація органів та тканин у філо- та онтогенезі «відібрала» для одних переважно клітинну форму, для інших – переважно або виключно внутрішньоклітинну, для третіх – однаково обидві форми регенерації (табл. 5). Переважна більшість тієї чи іншої форми регенерації у певних органах і тканинах визначається їх функціональним призначенням, структурно-функціональною спеціалізацією. Необхідність збереження цілісності покривів тіла пояснює, наприклад, переважання клітинної форми регенерації епітелію як шкіри, і слизових оболонок. Спеціалізована функція пірамідної клітини головного
мозку, як і м'язової клітини серця, виключає можливість поділу цих клітин та дозволяє зрозуміти необхідність відбору у філо- та онтогенезі внутрішньоклітинної регенерації як єдиної форми відновлення даного субстрату.
Таблиця 5.Форми регенерації в органах і тканинах ссавців (Саркісов Д.С., 1988)
Ці дані спростовують існуючі до недавнього часу уявлення про втрату деякими органами та тканинами ссавців здатності до регенерації, про «погано» і «добре» регенеруючі тканини людини, про те, що існує «закон зворотної залежності» між ступенем диференціювання тканин та здатністю їх до регенерації . В даний час встановлено, що в ході еволюції здатність до регенерації в деяких тканинах та органах не зникла, а прийняла форми (клітинну або внутрішньоклітинну), що відповідають їх структурній та функціональній своєрідності (Саркісов Д.С., 1977). Таким чином, всі тканини та органи мають здатність до регенерації, різні лише її форми залежно від структурно-функціональної спеціалізації тканини або органу.
МорфогенезРегенераторний процес складається з двох фаз - проліферації та диференціювання. Особливо добре ці фази виражені при формі клітинної регенерації. У фазу проліферації розмножуються молоді, недиференційовані клітини. Ці клітини називають камбіальними(Від лат. cambium- обмін, зміна), стовбуровими клітинамиі клітинами-попередниками.
Для кожної тканини характерні свої камбіальні клітини, які відрізняються ступенем проліферативної активності та спеціалізації, проте одна стовбурова клітина може бути родоначальником кількох видів
клітин (наприклад, стовбурова клітина кровотворної системи, лімфоїдної тканини, деякі клітинні представники сполучної тканини).
У фазу диференціювання молоді клітини дозрівають, відбувається їх структурно-функціональна спеціалізація. Та ж зміна гіперплазії ультраструктур їх диференціюванням (дозріванням) лежить в основі механізму внутрішньоклітинної регенерації.
Регулювання регенераторного процесу.Серед регуляторних механізмів регенерації розрізняють гуморальні, імунологічні, нервові, функціональні.
Гуморальні механізмиреалізуються як у клітинах пошкоджених органів і тканин (внутрішньотканинні та внутрішньоклітинні регулятори), так і за їх межами (гормони, поетини, медіатори, фактори росту та ін.). До гуморальних регуляторів відносять кейлони (Від грец. chalaino- послаблювати) - речовини, здатні пригнічувати поділ клітин та синтез ДНК; вони мають тканинну специфічність. Імунологічні механізмирегуляції пов'язані з "регенераційною інформацією", що переноситься лімфоцитами. У зв'язку з цим слід зауважити, що механізми імунологічного гомеостазу визначають структурний гомеостаз. Нервові механізмирегенераторних процесів пов'язані насамперед із трофічною функцією нервової системи, а функціональні механізми- з функціональним «запитом» органу, тканини, що розглядається як стимул до регенерації.
Розвиток регенераторного процесу багато в чому залежить від низки загальних та місцевих умов, чи факторів. До загальним слід віднести вік, конституцію, характер харчування, стан обміну та кровотворення, місцевим - стан іннервації, крово- та лімфообігу тканини, проліферативну активність її клітин, характер патологічного процесу.
Класифікація.Розрізняють три види регенерації: фізіологічну, репаративну та патологічну.
Фізіологічна регенераціявідбувається протягом усього життя і характеризується постійним оновленням клітин, волокнистих структур, основної речовини сполучної тканини. Немає таких структур, які б не зазнавали фізіологічної регенерації. Там, де домінує клітинна форма регенерації, має місце відновлення клітин. Так відбувається постійна зміна покривного епітелію шкіри та слизових оболонок, секреторного епітелію екзокринних залоз, клітин, що вистилають серозні та синовіальні оболонки, клітинних елементів сполучної тканини, еритроцитів, лейкоцитів та тромбоцитів крові тощо. У тканинах та органах, де клітинна форма регенерації втрачена, наприклад, у серці, головному мозку, відбувається оновлення внутрішньоклітинних структур. Поряд з оновленням клітин та субклітинних структур постійно відбувається біохімічна регенерація,тобто. поновлення молекулярного складу всіх компонентів тіла.
Репаративна чи відновна регенераціяспостерігається при різних патологічних процесах, що ведуть до пошкодження клітин і тканини.
ній. Механізми репаративної та фізіологічної регенерації єдині, репаративна регенерація – це посилена фізіологічна регенерація. Однак через те, що репаративна регенерація спонукається патологічними процесами, вона має якісні морфологічні відмінності від фізіологічної. Репаративна регенерація може бути повною та неповною.
Повна регенерація,або реституція,характеризується відшкодуванням дефекту тканиною, яка ідентична загиблої. Вона розвивається переважно у тканинах, де переважає клітинна регенерація.Так, у сполучній тканині, кістках, шкірі та слизових оболонках навіть відносно великі дефекти органу можуть шляхом поділу клітин заміщатися тканиною, ідентичною загиблої. При неповної регенерації,або субституції,дефект заміщається сполучною тканиною, рубцем. Субституція характерна для органів і тканин, в яких переважає внутрішньоклітинна форма регенерації, або вона поєднується з регенерацією клітин. Оскільки при регенерації відбувається відновлення структури, здатної до виконання спеціалізованої функції, сенс неповної регенерації над заміщенні дефекту рубцем, а компенсаторної гіперплазіїелементів спеціалізованої тканини, маса якої збільшується, тобто. відбувається гіпертрофіятканини.
При неповної регенерації,тобто. загоєнні тканини рубцем, виникає гіпертрофія як вираження регенераторного процесу, тому її називають регенераційної,у ній – біологічний сенс репаративної регенерації. Регенераторна гіпертрофія може здійснюватися двома шляхами - за допомогою гіперплазії клітин або гіперплазії та гіпертрофії клітинних ультраструктур, тобто. гіпертрофії клітин.
Відновлення вихідної маси органу та його функції за рахунок переважно гіперплазії клітинвідбувається при регенераційній гіпертрофії печінки, нирок, підшлункової залози, надниркових залоз, легень, селезінки та ін. Регенераційна гіпертрофія за рахунок гіперплазії клітинних ультраструктурй у міокарда, мозку, тобто. органів, де переважає внутрішньоклітинна форма регенерації. У міокарді, наприклад, по периферії рубця, який замістив інфаркт, розміри волокон м'язів значно збільшуються, тобто. вони гіпертрофуються у зв'язку з гіперплазією їх субклітинних елементів (рис. 81). Обидва шляхи регенераційної гіпертрофії не виключають одна одну, а, навпаки, нерідко поєднуються. Так, при регенераційній гіпертрофії печінки відбувається не тільки збільшення числа клітин у частині органу, що збереглася після пошкодження, а й гіпертрофія їх, обумовлена гіперплазією ультраструктур. Не можна виключити те, що у м'язі серця регенераційна гіпертрофія може протікати у вигляді гіпертрофії волокон, а й шляхом збільшення кількості складових їх м'язових клітин.
Відновлювальний період зазвичай не обмежується лише тим, що у пошкодженому органі розгортається репаративна регенерація. Якщо
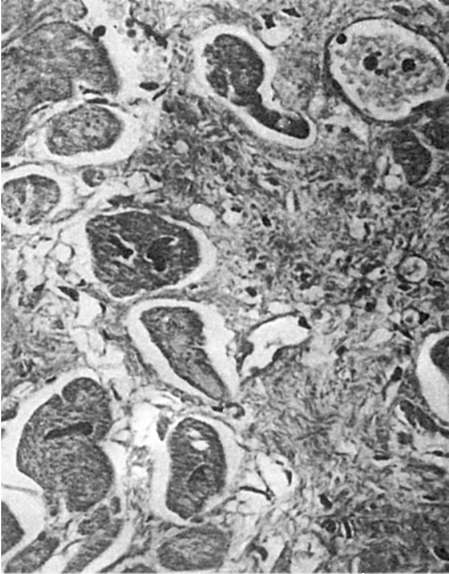 Мал. 81.Регенераційна гіпертрофія міокарда. По периферії рубця розташовані гіпертрофовані м'язові волокна
Мал. 81.Регенераційна гіпертрофія міокарда. По периферії рубця розташовані гіпертрофовані м'язові волокна
Вплив патогенного фактора припиняється до загибелі клітини, відбувається поступове відновлення пошкоджених органел. Отже, прояви репаративної реакції мають бути розширені за рахунок включення внутрішньоклітинних відновних процесів у дистрофічно змінених органах. Загальноприйнята думка про регенерацію лише як завершальний етап патологічного процесу маловиправдано. Репаративна регенерація не місцева, а загальна реакція організму, що охоплює різні органи, але реалізується повною мірою лише тому чи іншому їх.
Про патологічної регенерації говорять у тих випадках, коли внаслідок тих чи інших причин є збочення регенераторного процесу, порушення зміни фазпроліферації
та диференціювання. Патологічна регенерація проявляється у надмірному або недостатньому утворенні регенеруючої тканини (Гіпер-або гіпорегенерація),а також у перетворенні в ході регенерації одного виду тканини на іншу [метаплазія - див. Процеси пристосування (адаптації) та компенсації].Прикладами можуть бути гіперпродукція сполучної тканини з утворенням келоїда,надмірна регенерація периферичних нервів та надмірне утворення кісткової мозолі при зрощенні перелому, мляве загоєння ран та метаплазія епітелію в осередку хронічного запалення. Патологічна регенерація зазвичай розвивається при порушеннях загальнихі місцевих умов регенерації(порушення іннервації, білкове та вітамінне голодування, хронічне запалення тощо).
Регенерація окремих тканин та органів
Репаративна регенерація крові відрізняється від фізіологічної насамперед своєю більшою інтенсивністю. При цьому активний червоний кістковий мозок з'являється у довгих трубчастих кістках на місці жирового кісткового мозку (мієлоїдне перетворення жирового кісткового мозку). Жирові клітини витісняються острівцями кровотворної тканини, що росте, яка заповнює кістковомозковий канал і виглядає соковитою, темночервоною. Крім того, кровотворення починає відбуватися поза кістковим мозком. позакістномозкове,або екстрамедулярне, кровотворення.Оча-
гі екстрамедулярного (гетеротопічного) кровотворення в результаті виселення з кісткового мозку стовбурових клітин з'являються в багатьох органах і тканинах - селезінці, печінці, лімфатичних вузлах, слизових оболонках, жировій клітковині і т.д.
Регенерація крові може бути різко пригноблена (наприклад, при променевій хворобі, апластичній анемії, алейкії, агранулоцитозі) або перекручена (Наприклад, при злоякісній анемії, поліцитемії, лейкозі). У кров при цьому надходять незрілі, функціонально неповноцінні і формові елементи, що швидко руйнуються. У таких випадках говорять про патологічної регенерації крові
Репаративні можливості органів кровотворної та імунокомпетентної системи неоднозначні. Кістковий мозок має дуже високі пластичні властивості і може відновлюватися навіть при значних пошкодженнях. Лімфатичні вузли добре регенерують тільки в тих випадках, коли зберігаються зв'язки лімфатичних судин, що приносять і виносять, з навколишньою сполучною тканиною. Регенерація тканини селезінки при пошкодженні буває, як правило, неповною, загибла тканина заміщується рубцем.
Регенерація кровоносних та лімфатичних судинпротікає неоднозначно залежно від їхнього калібру.
Мікросудини мають більшу здатність регенерувати, ніж великі судини. Новоутворення мікросудин може відбуватися шляхом брунькування або аутогенно. При регенерації судин шляхом брунькування (рис. 82) в їх стінці з'являються бічні випинання за рахунок ендотеліальних клітин, що посилено діляться (ангіобласти). Утворюються тяжі з ендотелію, в яких виникають просвіти і в них надходить кров або лімфа з материнської судини. Інші елементи: судинної стінки утворюються за рахунок диференціювання ендотелію і оточуючих судин сполучнотканинних клітин, У судинну стінку вростають нервові волокна з передіснуючих нервів. Аутогенне новоутворення судин у тому, що у сполучної тканини з'являються осередки недиференційованих клітин. У цих осередках виникають щілини, у яких відкриваються передіснуючі капіляри і кров. Молоді клітини сполучної тканини, що диференціюються, утворюють ендотеліальну вистилку та інші елементи стінки судини.
 Мал. 82.Регенерація судин шляхом брунькування
Мал. 82.Регенерація судин шляхом брунькування
Великі судини не мають достатніх пластичних властивостей. Тому при пошкодженні їх стінки відновлюються лише структури внутрішньої оболонки, її ендотеліальна вистилка; елементи середньої та зовнішньої оболонок зазвичай заміщаються сполучною тканиною, що веде нерідко до звуження або облітерації просвіту судини.
Регенерація сполучної тканинипочинається з проліферації молодих мезенхімальних елементів та новоутворення мікросудин. Утворюється молода, багата на клітини і тонкостінні судини сполучна тканина, яка має характерний вигляд. Це - соковита темно-червона тканина з зернистою, ніби посипаною великими гранулами поверхнею, що стало підставою назвати її грануляційною тканиною.Гранули є виступаючими над поверхнею петлі новоутворених тонкостінних судин, які становлять основу грануляційної тканини. Між судинами багато недиференційованих лімфоцитоподібних клітин сполучної тканини, лейкоцитів, плазматичних клітин та лаброцитів (рис. 83). Надалі відбувається дозрівання грануляційної тканини, основу якої лежить диференціювання клітинних елементів, волокнистих структур, і навіть судин. Число гематогенних елементів зменшується, а фібробластів – збільшується. У зв'язку із синтезом фібробластами колагену у міжклітинних просторах утворюються аргірофільні(див. рис. 83), а потім і колагенові волокна.Синтез фібробластами глікозаміногліканів служить утворенню
основної речовини сполучної тканини. У міру дозрівання фібробластів кількість колагенових волокон збільшується, вони групуються у пучки; одночасно зменшується кількість судин, вони диференціюються в артерії та вени. Дозрівання грануляційної тканини завершується утворенням грубоволокниста рубцева тканина.
Новоутворення сполучної тканини відбувається не тільки при її пошкодженні, але і при неповній регенерації інших тканин, а також при організації (інкапсуляції), загоєнні ран, продуктивному запаленні.
Дозрівання грануляційної тканини може мати ті чи інші відхилення. Запалення, що розвивається в грануляційній тканині, призводить до затримки її дозрівання,
 Мал. 83.Грануляційна тканина. Між тонкостінними судинами багато недиференційованих клітин сполучної тканини та аргірофільних волокон. Імпрегнація сріблом
Мал. 83.Грануляційна тканина. Між тонкостінними судинами багато недиференційованих клітин сполучної тканини та аргірофільних волокон. Імпрегнація сріблом
а надмірна синтетична активність фібробластів - до надмірної освіти колагенових волокон з подальшим різко вираженим гіалінозом. У таких випадках виникає рубцева тканина у вигляді пухлиноподібного утворення синюшно-червоного кольору, що височить над поверхнею шкіри у вигляді келоїду.Келоїдні рубці утворюються після різних травматичних уражень шкіри, особливо опіків.
Регенерація жирової тканинивідбувається за рахунок новоутворення сполучнотканинних клітин, які перетворюються на жирові (адипозоцити) шляхом накопичення в цитоплазмі ліпідів. Жирові клітини складаються в часточки, між якими розташовуються сполучнотканинні прошарки з судинами та нервами. Регенерація жирової тканини може відбуватися також з залишків ядросодержащих цитоплазми жирових клітин.
Регенерація кісткової тканинипри переломі кісток значною мірою залежить від ступеня руйнування кістки, правильної репозиції кісткових уламків, місцевих умов (стан кровообігу, запалення тощо). При неускладненому кістковому переломі, коли кісткові уламки нерухомі, може відбуватися первинне кісткове зрощення(Рис. 84). Воно починається з вростання в область дефекту та гематоми між уламками кістки молодих мезенхімальних елементів та судин. Виникає так звана попередня сполучнотканинна мозоль,в якій відразу ж починається утворення кістки. Воно пов'язане з активацією та проліферацією остеобластіву зоні ушкодження, але насамперед у періостаті та ендостаті. В остеогенній фіброретикулярній тканині з'являються малозвапнілі кісткові балочки, кількість яких наростає.
Утворюється попередня кісткова мозоль.Надалі вона дозріває і перетворюється на зрілу пластинчасту кістку - так утворюється
 Мал. 84.Первинне кісткове зрощення. Інтермедіарна кісткова мозоль (показана стрілкою), що спаює уламки кістки (за Г.І. Лаврищевою)
Мал. 84.Первинне кісткове зрощення. Інтермедіарна кісткова мозоль (показана стрілкою), що спаює уламки кістки (за Г.І. Лаврищевою)
остаточна кісткова мозоль,яка за своєю будовою відрізняється від кісткової тканини лише безладним розташуванням кісткових перекладин. Після того, як кістка починає виконувати свою функцію і з'являється статичне навантаження, знову утворена тканина за допомогою остеокластів і остеобластів піддається перебудові, з'являється кістковий мозок, відновлюються васкуляризація та іннервація. При порушенні місцевих умов регенерації кістки (розлад кровообігу), рухливості уламків, великих діафізарних переломах відбувається вторинне кісткове зрощення(Рис. 85). Для цього виду кісткового зрощення характерно утворення між кістковими уламками спочатку хрящової тканини, на основі якої будується кісткова тканина. Тому при вторинному кістковому зрощенні говорять про попередньої кістково-хрящової мозолі,яка згодом перетворюється на зрілу кістку. Вторинне кісткове зрощення порівняно з первинним зустрічається значно частіше та займає більше часу.
При несприятливі умови регенерація кісткової тканини може бути порушена. При інфікуванні рани регенерація кістки затримується. Кісткові уламки, які при нормальному перебігу регенераторного процесу виконують функцію каркасу для новоствореної кісткової тканини, в умовах нагноєння рани підтримують запалення, що гальмує регенерацію. Іноді первинна кістково-хрящова мозоль не диференціюється у кісткову. У цих випадках кінці зламаної кістки залишаються рухливими, утворюється хибний суглоб.Надмірна продукція кісткової тканини в ході регенерації призводить до появи кісткових виростів. екзостозів.
Регенерація хрящової тканинина відміну кісткової відбувається зазвичай неповно. Лише невеликі дефекти її можуть заміщати новоствореною тканиною за рахунок камбіальних елементів надхрящниці. хондробластів.Ці клітини створюють основну речовину хряща, потім перетворюються на зрілі клітини хряща. Великі дефекти хряща заміняються рубцевою тканиною.
Регенерація м'язової тканиниїї можливості та форми різні залежно від виду цієї тканини. Гладкі м'щі, клітини яких мають здатність до мітозу і амітозу, при незначних дефектах можуть регенерувати досить повно. Значні ділянки пошкодження гладких м'язів заміщаються рубцем, при цьому м'язові волокна, що збереглися, піддаються гіпертрофії. Новоутворення гладких м'язових волокон може відбуватися шляхом перетворення (метаплазії) елементів сполучної тканини. Так утворюються пучки гладких м'язових волокон в плевральних спайках, в організації, що піддаються тромбах, в судинах при їх диференціюванні.
Поперечносмугасті м'язи регенерують лише за збереження сарколеммы. Усередині трубок із сарколеми здійснюється регенерація її органел, внаслідок чого з'являються клітини, які називаються міобластами.Вони витягуються, кількість ядер у них збільшується, у саркоплазмі
 Мал. 85.Вторинне кісткове зрощення (за Г.І. Лаврищевою):
Мал. 85.Вторинне кісткове зрощення (за Г.І. Лаврищевою):
а - кістково-хрящова періостальна мозоль; ділянку кісткової тканини серед хрящової (мікроскопічна картина); б - періостальна кістково-хрящова мозоль (гістотопом через 2 міс після операції): 1 - кісткова частина; 2 – хрящова частина; 3 - уламки кістки; в - періостальна мозоль, що спаює зміщені уламки кістки
диференціюються міофібрили, і трубки сарколеми перетворюються на поперечносмугасті м'язові волокна. Регенерація скелетних м'язів може бути пов'язана і з клітинами-сателітами,які розташовуються під сарколемою, тобто. всередині м'язового волокна, і є камбіальними.У разі травми клітини-сателіти починають посилено ділитися, потім диференціюються і забезпечують відновлення м'язових волокон. Якщо при пошкодженні м'яза цілість волокон порушується, то на кінцях їх розривів виникають колбоподібні вибухання, які містять велику кількість ядер і називаються м'язовими бруньками.У цьому відновлення безперервності волокон немає. Місце розриву заповнюється грануляційною тканиною, що перетворюється на рубець (М'язова мозоль).Регенерація м'язи серця при її пошкодженні, як і при пошкодженні поперечно мускулатури, закінчується рубцюванням дефекту. Однак у м'язових волокнах, що збереглися, відбувається інтенсивна гіперплазія ультраструктур, що веде до гіпертрофії волокон і відновлення функції органу (див. рис. 81).
Регенерація епітеліюздійснюється в більшості випадків досить повно, так як він має високу регенераторну здатність. Особливо добре регенерує покривний епітелій. Відновлення багатошарового плоского ороговіючого епітелію можливо навіть за досить великих дефектів шкіри. При регенерації епідермісу у краях дефекту відбувається посилене розмноження клітин зародкового (камбіального), росткового (мальпігієвого) шару. Епітеліальні клітини, що утворюються, спочатку покривають дефект одним шаром. Надалі пласт епітелію стає багатошаровим, клітини його диференціюються, і він набуває всіх ознак епідермісу, що включає ростковий, зернистий блискучий (на підошвах і долонній поверхні кистей) і роговий шари. При порушенні регенерації епітелію шкіри утворюються виразки, що не гояться, нерідко з розростанням в їх краях атипового епітелію, що може послужити основою для розвитку раку шкіри.
Покривний епітелій слизових оболонок (багатошаровий плоский неороговуючий, перехідний, одношаровий призматичний і багатоядерний миготливий) регенерує таким же чином, як і багатошаровий плоский ороговіючий. Дефект слизової оболонки відновлюється за рахунок проліферації клітин, що вистилають крипти та вивідні протоки залоз. Недиференційовані сплощені клітини епітелію спочатку покривають дефект тонким шаром (мал. 86), потім клітини набувають форми, властиву клітинним структурам відповідної епітеліальної вистилки. Паралельно частково або повністю відновлюються залози слизової оболонки (наприклад, трубчасті залози кишки, залози ендометрію).
Регенерація мезотеліюочеревини, плеври і навколосерцевої сумки здійснюється шляхом розподілу клітин, що збереглися. На поверхні дефекту з'являються порівняно великі кубічні клітини, які потім ущільнюються. При невеликих дефектах мезотеліальна вистилка відновлюється швидко та повно.
Важливе значення для відновлення покривного епітелію і мезотелію має стан сполучної тканини, що підлягає, так як епітелізація будь-якого дефекту можлива лише після заповнення його грануляційною тканиною.
Регенерація спеціалізованого епітелію органів(печінки, підшлункової залози, нирок, залоз внутрішньої секреції, легеневих альвеол) здійснюється за типом регенераційної гіпертрофії:в ділянках ушкодження тканина заміщується рубцем, а по периферії його відбуваються гіперплазія та гіпертрофія клітин паренхіми. У печінки ділянка некрозу завжди піддається рубцюванню, однак у решті органу відбувається інтенсивне новоутворення клітин, а також гіперплазія внутрішньоклітинних стуктур, що супроводжується їх гіпертрофією. В результаті цього вихідна маса та функція органу швидко відновлюються. Регенераторні можливості печінки майже безмежні. У підшлунковій залозі регенераторні процеси добре виражені як в екзокринних відділах, так і в панкреатичних острівцях, причому епітелій екзокринних залоз стає джерелом відновлення острівців. У нирках при некрозі епітелію канальців відбувається розмноження нефроцитів, що збереглися, і відновлення канальців, проте лише при збереженні тубулярної базальної мембрани. За її руйнації (тубулорексис) епітелій не відновлюється і каналець заміщається сполучною тканиною. Чи не відновлюється загиблий канальцевий епітелій і в тому випадку, коли одночасно з канальцем гине судинний клубочок. При цьому на місці загиблого нефрону розростається рубцева сполучна тканина, а навколишні нефрони піддаються регенераційній гіпертрофії. У залозах внутрішньої секреції відновлювальні процеси також представлені неповною регенерацією. У легкому після видалення окремих часток у частині, що залишилася, відбувається гіпертрофія і гіперплазія тканинних елементів. Регенерація спеціалізованого епітелію органів може протікати атипово, що веде до розростання сполучної тканини, структурної перебудови та деформації органів; у таких випадках говорять про цирозі (Цироз печінки, нефроцироз, пневмоцироз).
Регенерація різних відділів нервової системивідбувається неоднозначно. У головному і спинному мозку новоутворення гангліозних клітин не про-
 Мал. 86.Регенерація епітелію у дні хронічної виразки шлунка
Мал. 86.Регенерація епітелію у дні хронічної виразки шлунка
виходить і при руйнуванні їх відновлення функції можливе лише за рахунок внутрішньоклітинної регенерації клітин, що збереглися. Невроглії, особливо мікроглії, властива клітинна форма регенерації, тому дефекти тканини головного та спинного мозку зазвичай заповнюються проліферуючими клітинами невроглії – виникають так звані гліальні (гліозні) рубці. При пошкодженні вегетативних вузлів поряд з гіперплазією ультраструктур клітин відбувається та їх новоутворення. При порушенні цілості периферичного нерва регенерація відбувається за рахунок центрального відрізка, що зберіг зв'язок з клітиною, тоді як периферичний відрізок гине. Клітини, що розмножуються, шваннівської оболонки загиблого периферичного відрізка нерва розташовуються вздовж нього і утворюють футляр - так званий бюнгнерівський тяж, в який вростають регенеруючі осьові циліндри з проксимального відрізка. Регенерація нервових волокон завершується їх мієлінізацією та відновленням нервових закінчень. Регенераційна гіперплазія рецепторів, перицелюлярних синаптичних приладів та ефекторів іноді супроводжується гіпертрофією їх кінцевих апаратів. Якщо регенерація нерва з тих чи інших причин порушується (значне розбіжність частин нерва, розвиток запального процесу), то місці його перерви утворюється рубець, у якому безладно розташовуються регенерировавшие осьові циліндри проксимального відрізка нерва. Аналогічні розростання виникають на кінцях перерізаних нервів у культі кінцівки після її ампутації. Такі розростання, утворені нервовими волокнами та фіброзною тканиною, називаються ампутаційними невромами
Загоєння ран
Загоєння ран протікає за законами репаративної регенерації. Темпи загоєння ран, його результати залежать від ступеня та глибини ранового ушкодження, структурних особливостей органу, загального стану організму, методів лікування, що застосовуються. За І.В. Давидовському виділяють такі види загоєння ран: 1) безпосереднє закриття дефекту епітеліального покриву; 2) загоєння під струпом; 3) загоєння рани первинним натягом; 4) загоєння рани вторинним натягом, чи загоєння рани через нагноєння.
Безпосереднє закриття дефекту епітеліального покриву- це найпростіше загоєння, що полягає в наповзанні епітелію нд поверхневий дефект і закриття його епітеліальним шаром. Спостерігається на рогівці, слизових оболонках загоєння під струпомстосується дрібних дефектів, на поверхні яких швидко виникає скоринка, що підсихає (струп) з згорнутої крові і лімфи; епідерміс відновлюється під скоринкою, яка відпадає через 3-5 діб після поранення.
Загоєння первинним натягом (per rimamm intentionem)спостерігається в ранах з пошкодженням не тільки шкіри, але і тканини, що підлягає,
причому краї рани рівні. Рана заповнюється згортками крові, що вилилася, що оберігає краї рани від дегідратації та інфекції. Під впливом протеолітичних ферментів неітрофілів відбувається частковий лізис згортка крові, тканинного детриту. Нейтрофіли гинуть, на зміну їм приходять макрофаги, які фагоцитують еритроцити, залишки пошкодженої тканини; у краях рани виявляється гемосидерин. Частина вмісту рани видаляється в перший день поранення разом з ексудатом самостійно або при обробці рани - первинне очищення. На 2-3 добу в краях рани з'являються фібробласти і новоутворені капіляри, що ростуть назустріч один одному, з'являється грануляційна тканина,пласт якої при первинному натягу не досягає великих розмірів. До 10-15-ї доби вона повністю дозріває, рановий дефект епітелізується і рана гоїться ніжним рубчиком. У хірургічній рані загоєння первинним натягом прискорюється у зв'язку з тим, що її краї стягуються нитками шовку або кетгуту, навколо яких скупчуються гігантські клітини сторонніх тіл, що їх розсмоктують, не заважають загоєнню.
Загоєння вторинним натягом (per secundam intentionem),або загоєння через нагноєння (або загоєння за допомогою гранулювання - per granulationem),спостерігається зазвичай при великих пораненнях, що супроводжуються розмозженням і омертвінням тканин, проникненні в рану сторонніх тіл, мікробів. На місці рани виникають крововиливи, травматичний набряк країв рани, швидко з'являються ознаки демаркаційного. гнійного запаленняна кордоні з відмерлою тканиною, розплавлення некротичних мас. Протягом перших 5-6 діб відбувається відторгнення некротичних мас - вторинне очищення рани і в краях рани починає розвиватися грануляційна тканина. Грануляційна тканина,виконує рану, складається з 6 шарів, що переходять один в одного (Анічков Н.Н., 1951): поверхневий лейкоцитарно-некротичний шар; поверхневий шар судинних петель, шар вертикальних судин, шар, що дозріває, шар горизонтально розташованих фібробластів, фіброзний шар. Дозрівання грануляційної тканини при загоєнні рани вторинним натягом супроводжується регенерацією епітелію. Однак при цьому виді загоєння рани дома її завжди утворюється рубець.
Регенерація- відновлення організмом втрачених або пошкоджених органів та тканин, а також відновлення цілого організму з його частини. Більшою
ступеня властива рослинам і безхребетним тваринам, меншою - хребетним. Регенерацію можна викликати
експериментально.
Регенераціяспрямована на відновлення пошкоджених структурних елементів та регенераційні процеси можуть
здійснюватись на різних рівнях:
а) молекулярний
б) субклітинний
в) клітинний – розмноження клітин мітозом та амітотичним шляхом
г) тканинний
д) органний.
Види регенерації:
7. Фізіологічна -забезпечує функціонування органів та систем у звичайних умовах. У всіх органах відбувається фізіологічна регенерація, але в якихось більше, в інших – менше.
2. Репаративна(відновна) - виникає у зв'язку з патологічним процесом, що призводить до пошкодження тканини (це посилена фізіологічна регенерація)
а) повна регенерація (реституція) - на місці пошкодження тканини виникає така сама тканина
б) неповна регенерація (субституція) – на місці загиблої тканини виникає сполучна тканина. Наприклад, серце при інфаркті міокарда відбувається некроз, який заміщається сполучною тканиною.
Сенс неповної регенерації:навколо сполучної тканини виникає регенераційна гіпертрофія, яка і
забезпечує збереження функції ушкодженого органу.
Регенераційна гіпертрофіяздійснюється за рахунок:
а) гіперплазії клітин (надлишкова освіта)
б) гіпертрофії клітин (збільшення органу в обсязі та масі).
Регенераційна гіпертрофія у міокарді здійснюється за рахунок гіперплазії внутрішньоклітинних структур.
Форми регенерації.
1. Клітинна – відбувається розмноження клітин мітотичним та амітотичним шляхом. Вона існує в кістковій тканині, епідермісі, слизовій оболонці ШКТ, слизовій оболонці дихальних шляхів, слизовій сечостатевої системи, ендотелій, мезотелій, пухка сполучна тканина, кровотворна система. У цих органах і тканинах виникає повна регенерація (точно така сама тканина).
2. Внутрішньоклітинна – відбувається гіперплазія внутрішньоклітинних структур. Міокард, скелетні м'язи (переважно), гангліозні клітини ЦНС (виключно).
3. Клітинні та внутрішньоклітинні форми. Печінка, нирки, легені, гладкі м'язи, вегетативна нервова система, підшлункова залоза, ендокринна система. Зазвичай виникає неповна регенерація.
Регенерація сполучної тканини.
Етапи:
1. Утворення грануляційної тканини. Поступово йде витіснення судин та клітин з утворенням волокон. Фібробласти – фіброцити, які продукують волокна.
2. Утворення зрілої сполучної тканини. Регенерація крові
1. Фізіологічна регенерація. У кістковому мозку.
2. Репаративна регенерація. Виникає при анеміях, лейкопеніях, тромбоцитопеніях. З'являються екстрамедулярні осередки кровотворення (у печінці, селезінці, лімфатичних вузлах, жовтий кістковий мозок бере участь у кровотворенні).
3. Патологічна регенерація. При променевій хворобі, лейкозах. В органах кровотворення утворюються незрілі
кровотворні елементи (владні клітини)
Запитання 16
ГОМЕОСТАЗ.
Гомеостаз - Підтримка сталості внутрішнього середовища організму в умовах зовнішнього середовища, що безперервно змінюються. Т.к. організм - багаторівневий об'єкт, що саморегулюється, його можна розглядати з точки зору кібернетики. Тоді організм - складна багаторівнева саморегулююча система з безліччю змінних.
Змінні входи:
Причина;
Роздратування.
Змінні виходи:
реакція;
Слідство.
Причина відхилення від норми реакції в організмі. Вирішальна роль належить зворотний зв'язок. Існує позитивний і негативний зворотний зв'язок.
Негативний зворотний зв'язокзменшує дію вхідного сигналу на вихідний. Позитивний зворотний зв'язокзбільшує дію вхідного сигналу вихідний ефект дії.
Живий організм – ультрастабільна система, яка здійснює пошук найбільш оптимального сталого стану, що забезпечується адаптаціями.
Питання 18:
ПРОБЛЕМИ ТРАНСПЛАНТАЦІЇ.
Трансплантація-пересадка тканин та органів.
Трансплантація у тварин та людини - приживлення органів чи ділянок окремих тканин для заміщення дефектів, стимулювання регенерації, при косметичних операціях, а також з метою експерименту та тканинної терапії.
Аутотрансплантація-пересадка тканин у межах одного організму Алотрансплантація-пересадка між організмами одного виду. Ксенотрансплантація - пересадка між різними видами.
Запитання 19
Хронобіологія- розділ біології, що вивчає біологічні ритми, перебіг різних біологічних процесів
(переважно циклічних) у часі.
Біологічні ритми- (біоритми), циклічні коливання інтенсивності та характеру біологічних процесів та явищ. Одні біологічні ритми щодо самостійні (напр., частота скорочень серця, дихання), інші пов'язані з пристосуванням організмів до геофізичних циклів - добовим (напр., коливання інтенсивності поділу клітин, обміну речовин, рухової активності тварин), припливним (напр., біологічні процеси в організмів, пов'язані з рівнем морських припливів), річним (зміна чисельності та активності тварин, зростання та розвитку рослин та ін.). Наука про біологічні ритми – хронобіологія.
Запитання 20
Філогенез скелета
Скелет риб складається з черепа, хребта, скелета непарних, парних плавців та його поясів. У тулубному відділі до поперечних відростків тіла причленовуються ребра. Хребці зчленовуються один з одним за допомогою суглобових відростків, забезпечуючи вигин переважно у горизонтальній площині.
Скелет земноводних, як і у всіх хребетних, складається з черепа, хребта, скелета кінцівок та їх поясів. Череп майже суцільно хрящовий (рис. 11.20). Він рухливо зчленований з хребтом. Хребет містить дев'ять хребців, об'єднаних у три відділи: шийний (1 хребець), тулубовий (7 хребців), крижовий (1 хребець), проте хвостові хребці зрослися, утворивши єдину кісточку - уростиль. Ребра відсутні. Плечовий пояс включає типові для наземних хребетних кісток: парні лопатки, воронячі кістки (коракоїди), ключиці та непарну грудину. Він має вигляд півкільця, що лежить у товщі тулубової мускулатури, тобто не з'єднаний з хребтом. Тазовий пояс утворений двома тазовими кістками, утвореними трьома парами клубових, сідничних і лобкових кісток, що зрослися між собою. Довгі клубові кістки приєднані до поперечних відростків крижового хребця. Скелет вільних кінцівок побудований на кшталт системи многочленных важелів, рухомо з'єднаних кулястими суглобами. У складі передньої кінцівки. виділяють плече, передпліччя та кисть.
Тіло ящірки поділено на голову, тулуб і хвіст. У тулубному відділі добре виражена шия. Все тіло вкрите роговими лусками, а голова та черево – великими щитками. Кінцівки ящірки добре розвинені та озброєні п'ятьма пальцями з пазурами. Плечові та стегнові кістки розташовані паралельно поверхні землі, внаслідок чого тіло провисає і стосується землі (звідси і назва класу). Шийний відділ хребта складається з восьми хребців, перший їх рухомо з'єднаний як із черепом, і з другим хребцем, що забезпечує головному відділу більшу свободу рухів. Хребці попереково-грудного відділу несуть ребра, частина яких з'єднана з грудиною, у результаті утворюється грудна клітина. Хрестові хребці забезпечують міцніше, ніж у земноводних, з'єднання з кістками тазу.
Скелет ссавців за будовою в основному подібний до скелета наземних хребетних, проте є деякі відмінності: число шийних хребців постійно і дорівнює семи, череп більш об'ємний, що пов'язано з великими розмірами головного мозку. Кістки черепа зростаються досить пізно, що забезпечує можливість збільшення головного мозку зі зростанням тварини. Кінцівки ссавців побудовані за п'ятипалим типом, характерним для наземних хребетних.
Запитання 21
ФІЛОГЕНЕЗ КРОВЕНОСНОЇ СИСТЕМИ
Кровоносна система риб заткнута. Серце двокамерне, що складається з передсердя та шлуночка. Венозна кров із шлуночка серця надходить у черевну аорту, що несе її до зябер, де вона збагачується киснем і звільняється від вуглекислого газу. Відтікає від зябер артеріальна кров збирається в спинну аорту, розташовану вздовж тіла під хребтом. Від спинної аорти до різних органів риби відходять численні артерії. Вони артерії розпадаються на мережу найтонших, капілярів, через стінки яких кров віддає кисень і збагачується вуглекислим газом. Венозна кров збирається у вени і з них надходить у передсердя, та якщо з нього шлуночок. Отже, риби мають одне коло кровообігу.
Кровоносна система земноводних представлена трикамерним серцем, що складається з двох передсердь і шлуночка, і двох кіл кровообігу – великого (тулубового) та малого (легеневого). Мале коло кровообігу починається у шлуночку, включає судини легень і завершується у лівому передсерді. Велике коло починається також у шлуночку. Кров, пройшовши судинами всього тіла, повертається у праве передсердя. Таким чином, у ліве передсердя потрапляє артеріальна кров з легенів, а праве - венозна кров з усього тіла. У праве передсердя потрапляє і артеріальна кров, яка відтікає від шкіри. Так, завдяки появі легеневого кола кровообігу в серце земноводних потрапляє і артеріальна кров. Незважаючи на те, що в шлуночок надходить артеріальна та венозна кров, повного перемішування крові не відбувається завдяки наявності кишень і неповних перегородок. Завдяки їм при виході зі шлуночка артеріальна кров по сонних артеріях надходить у головний відділ, венозна - у легені та шкіру, а змішана - у решту органів тіла. Таким чином, у земноводних немає повного поділу крові у шлуночку, тому інтенсивність життєвих процесів невисока, а температура тіла непостійна.
Серце у плазунів трикамерне, проте повного змішування артеріальної та венозної крові не відбувається через наявність у ньому неповної поздовжньої перегородки. Три судини, що відходять від різних частин шлуночка - легенева артерія, ліва і права дуги аорти - несуть венозну кров до легень, артеріальну - до голови і передніх кінцівок, а до інших частин - змішану з переважанням артеріальної. Таке кровозабезпечення, а також мала здатність до терморегуляції призводять до того, що
температура тіла плазунів залежить від температурних умов довкілля.
Високий рівень життєдіяльності птахів зумовлений більш досконалою системою кровообігу, порівняно з тваринами попередніх класів. У них відбувся повний поділ артеріального та венозного потоків крові. Це з тим, що серце птахів чотирикамерне і повністю поділено на ліву - артеріальну, і праву - венозну, частини. Дуга аорти лише одна (права) і відходить від лівого шлуночка. У ній тече чиста артеріальна кров, що забезпечує всі тканини та органи тіла. Від правого шлуночка відходить легенева артерія, що несе в легені венозну кров. Кров швидко рухається судинами, газообмін відбувається інтенсивно, виділяється багато тепла. Кровоносна система ссавців важливих відмінностей від такої у птахів немає, На відміну від птахів, у ссавців від лівого шлуночка відходить ліва дуга аорти.
Запитання 22
РОЗВИТОК АРТЕРІАЛЬНИХ ДУГ
Артеріальні дуги, дуги аорти, кровоносні судини, що закладаються у зародків хребетних у вигляді 6-7 (у круглоротих до 15) парних бічних стволів, що відходять від черевної аорти. А. д. проходять по міжжаберним перегородкам на спинний бік глотки і, зливаючись, утворюють спинну аорту. Перші дві пари артеріальних дуг зазвичай рано редукуються, у риб і личинок земноводних вони зберігаються у вигляді невеликих судин. Інші 4-5 пар артеріальних дуг стають зябровими судинами. У наземних хребетних із третьої пари артеріальних дуг утворюються сонні артерії, із шостої - легеневі. У хвостатих земноводних зазвичай 4 і 5 пари артеріальних дуг утворюють стовбури або коріння аорти, що зливаються в спинну аорту. У безхвостих земноводних і плазунів дуги аорти виникають тільки з 4-ї пари артеріальних дуг, а 5-та редукується. У птахів і ссавців редуковані 5-та половина 4-ї артеріальних дуг, у птахів аортою стає її права половина, у ссавців - ліва. Іноді у дорослих особин зберігаються зародкові судини, що з'єднують дуги аорти з сонними (сонні протоки) або з легеневими (боталові протоки) артеріями.
Запитання 23
Дихальна система.
Більшість тварин – аероби. Дифузія газів з атмосфери у вигляді водного розчину здійснюється при диханні. Елементи шкірного та водного дихання зберігаються навіть у вищих хребетних тварин. У ході еволюції у тварин виникли різноманітні дихальні пристосування – похідні шкіри та травної трубки. Зябра та легені – похідні глотки.
ФІЛОГЕНЕЗ ОРГАНІВ ДИХАННЯ
Органи дихання - зябра - розташовані на верхній стороні чотирьох зябрових дуг у вигляді яскраво-червоних пелюсток. Вода потрапляє в рот риби, проціджується через зяброві щілини, омиваючи зябра, і виводиться назовні з-під зябрової кришки. Газообмін здійснюється в численних зябрових капілярах, кров у яких тече назустріч воді, що омиває зябра.
Дихають жаби легкими та шкірою. Легкі є парні порожнисті мішки з пористої внутрішньої поверхнею, пронизаною мережею кровоносних капілярів, де відбувається газообмін. Механізм дихання у земноводних недосконалий, нагнітального типу. Тварина набирає повітря в порожнину ротоглоточную, для чого опускає дно ротової порожнини і відкриває ніздрі. Потім ніздрі закриваються клапанами, дно ротової порожнини піднімається, повітря нагнітається в легкі. Видалення повітря із легень відбувається завдяки скороченню грудних м'язів. Поверхня легень у земноводних невелика, менша за поверхню шкіри.
Органи дихання-легкі (рептилії). Їх стінки мають пористу будову, що значною мірою. збільшує поверхню. Шкірне дихання відсутнє. Вентиляція легень інтенсивніша, ніж у земноводних, і пов'язана зі зміною об'єму грудної клітини. Дихальні шляхи -трахея, бронхи - захищають легені від сушіння і охолоджувального впливу повітря, що надходить ззовні.
Легкі птахи є щільними губчастими тілами. Бронхи, увійшовши в легені, сильно в них розгалужуються до найтонших, сліпо замкнутих бронхіол, обплутаних мережею капілярів, де
та відбувається газообмін. Частина великих бронхів, не розгалужуючись, виходить межі легень і розширюється у величезні тонкостінні повітряні мішки, обсяг яких у багато разів перевищує обсяг легень (рис. 11.23). Повітряні мішки розташовані між різними внутрішніми органами, які відгалуження проходять між м'язами, під шкіру й у порожнини кісток.
Дихають ссавці легень, які мають альвеолярну структуру, завдяки якій дихальна поверхня перевершує поверхню тіла в 50 разів і більше. Механізм дихання обумовлений зміною об'єму грудної клітки за рахунок руху ребер і властивого ссавцям особливого м'яза - діафрагми.
Запитання 24
ФІЛОГЕНЕЗ ГОЛОВНОГО МОЗКУ
Центральна нервова система риб складається з головного та спинного мозку. Головний мозок у риб, як у всіх хребетних, представлений п'ятьма відділами: переднім, проміжним, середнім, мозочком і довгастим мозком. Від переднього мозку відходять добре розвинені нюхові частки. Найбільшого розвитку досягає середній мозок, який здійснює аналіз зорових сприйняттів, а також мозочок, що регулює координацію рухів та збереження рівноваги.
Головний мозок земноводних має ті п'ять відділів, як і мозок риб. Однак відрізняється від нього великим розвитком переднього мозку, який у земноводних розділений на дві півкулі. Мозок недорозвинений у зв'язку з малою рухливістю і однооб. різним характером рухів земноводних.
Головний мозок плазунів у порівнянні з таким земноводних має краще розвинені мозочки і великі півкулі переднього мозку, поверхня якого має зачатки кори. Це зумовлює різноманітні і складніші форми пристосувальної поведінки.
Головний мозок птахів відрізняється від мозку плазунів великими розмірами півкуль переднього мозку і мозочка.
Головний мозок ссавців має відносно великі розміри через збільшення обсягу півкуль переднього мозку та мозочка. Розвиток переднього мозку відбувається за рахунок розростання його даху – мозкового склепіння, або кори мозку.
Запитання 25
ФІЛОГЕНЕЗ ВИДІЛЮВАЛЬНОЇ І ПІДЛОГОВИЙ СИСТЕМ
Органами виділення риб служать парні стрічковоподібні тулубові нирки, розташовані в порожнині тіла під хребтом. Вони втратили зв'язок із порожниною тіла та видаляють шкідливі продукти життєдіяльності, відфільтровуючи їх із крові. У прісноводних риб кінцевим продуктом білкового обміну є отруйний аміак. Він розчиняється великою кількістю води і тому риби виділяють багато рідкої сечі. Виведена з сечею вода легко, заповнюється за рахунок її постійного надходження через шкіру, зябра та з їжею. У морських риб кінцевим продуктом азотистого обміну служить менш отруйна сечовина, виведення якої потребує меншої кількості води. Сеча, що утворилася в нирках, по парних сечоводах відтікає в сечовий міхур, звідки виводиться назовні через видільний отвір. Парні статеві залози - яєчники та сім'яники - мають вивідні протоки. Запліднення у більшості риб зовнішнє і відбувається у воді
Органи виділення земноводних, як і риб, представлені тулубними нирками. Однак, на відміну від риб, вони мають вигляд сплощених компактних тіл, що лежать з боків.
крижового хребця. У нирках є клубочки, які відфільтровують із крові шкідливі продукти розпаду (переважно сечовину) і водночас важливі для організму речовини (цукри, вітаміни та інших.). Під час стоку по ниркових канальцях корисні організму речовини всмоктуються назад у кров, а сеча надходить по двох сечоводів у клоаку і звідти сечовий міхур. Після заповнення сечового міхура його м'язові стінки скорочуються, сеча виводиться в клоаку і викидається назовні. Втрати води з організму земноводних із сечею, як і в риб, поповнюються надходженням її через шкіру. Статеві залози парні. Парні яйцеводи впадають у клоаку, а сім'явивідні канали - в сечоводи.
Органи плазунів виділення представлені тазовими нирками, у яких загальна фільтраційна площа клубочків невелика, тоді як довжина канальців значна. Це сприяє інтенсивному зворотному всмоктуванню відфільтрованої клубочками води капіляри крові. Отже, виділення продуктів життєдіяльності у плазунів відбувається з мінімальними втратами води. У них, як і у наземних членистоногих, кінцевим продуктом виділення є сечова кислота, яка вимагає виведення з організму невеликої кількості води. Сеча по сечоводниках збирається в клоаку, а з неї в сечовий міхур, з якого у вигляді суспензії дрібних кристалів виводиться назовні.
Виділення ссавців. Тазові нирки ссавців подібні до будови такими птахів. Сеча з великим вмістом сечовини відтікає від нирок по сечоводів у сечовий міхур, а з нього виходить назовні.
Запитання 26
Філогенез покривів тіла:
Основні напрямки еволюції покровів хордових:
1) диференціювання на два шари: зовнішній – епідерміс, внутрішній – дерму та збільшення товщини дерми;
1) від одношарового епідермісу до багатошарового;
2) диференціювання дерми на 2 шари - сосочковий та сітчастий:
3) поява підшкірно-жирової клітковини та вдосконалення механізмів терморегуляції;
4) від одноклітинних залоз до багатоклітинних;
5) диференціювання різних похідних шкіри.
У нижчих хордових (ланцетник)епідерміс одношаровий, циліндричний, має залізисті клітини, що виділяють слиз. Дерма (коріум) представлена тонким шаром неоформленої сполучної тканини.
У нижчих хребетних епідерміс стає багатошаровим. Нижній шар - паростковий (базальний), клітини його діляться і заповнюють клітини вищележачих шарів. Дерма має правильно розташовані волокна, судини та нерви.
Похідними шкіри є: одноклітинні (у круглоротих риб) та багатоклітинні (у земноводних) слизові залози; луска: а) плакоїдна у хрящових риб, у розвитку якої беруть участь епідерміс та дерма; б) кісткова у кісткових риб, що розвивається за рахунок дерми.
Плакоїдна луска зовні покрита шаром емалі (ектодермального походження), під яким знаходяться дентин та пульпа (мезодермального походження). Луска і слиз виконують захисну функцію.
У земноводних шкіра тонка гладка, без луски. Шкіра містить велику кількість багатоклітинних слизових залоз, секрет яких зволожує покриви і має бактерицидні властивості. Шкіра бере участь у газообміні.
У вищих хребетних у зв'язку з виходом на сушу епідерміс стає сухим, має роговий шар.
У рептилійрозвиваються рогові луски, відсутні шкірні залози.
У ссавців:добре розвинені епідерміс та дерма, з'являєтьсяпідшкірно-жирова клітковина.
Запитання 27
ФІЛОГЕНЕЗ ХАРЧОВОЇ СИСТЕМИ.
Харчуються риби різноманітною їжею. Харчова спеціалізація відбивається на будову органів травлення. Рот веде в порожнину рота, в якій зазвичай є численні зуби, розташовані на щелепних, піднебінних та інших кістках. Слинні залози відсутні. З ротової порожнини їжа проходить у глотку, пробиту зябровими щілинами, і по стравоходу потрапляє в шлунок, залози якого рясно виділяють травні соки. У деяких риб (корпові та ряд інших) шлунка немає і їжа надходить відразу в тонкий кишечник, де під впливом комплексу ферментів, що виділяються залозами самого кишечника, печінки та підшлункової залози, відбувається розщеплення їжі та всмоктування розчинених поживних речовин. Диференціювання травної системи земноводних залишилося приблизно на тому ж рівні, що й у їхніх предків – риб. Загальна ротоглоточная порожнина перетворюється на короткий стравохід, його розташований слабо відокремлений шлунок, перехідний без різкої межі в кишечник. Кишечник закінчується прямою кишкою, що переходить у клоаку. Протоки травних залоз - печінки та підшлункової залози - впадають у дванадцятипалу кишку. У ротоглоточную порожнину відкриваються протоки відсутніх у риб слинних залоз, що змочують ротову порожнину та їжу. З наземним способом життя пов'язана поява у ротовій порожнині справжньої мови – основного органу видобутку їжі.
У травній системі плазунів краще, ніж земноводних, виражена диференціювання на відділи. Захоплення їжі проводиться щелепами, які мають утримання видобутку зуби. Ротова порожнина краще, ніж у земноводних, відмежована від горлянки. На дні ротової порожнини розташований рухливий, роздвоєний на кінці язик. Їжа змочується слиною, що полегшує її заковтування. Стравохід у зв'язку з розвитком шиї довгий. Відмежований від стравоходу шлунок має мускулисті стінки. На межі тонкої та товстої кишок є сліпа кишка. Протоки печінки та підшлункової
залози відкриваються у дванадцятипалу кишку. Час перетравлення їжі залежить від температури тіла плазунів.
Травна система ссавців. Зуби сидять у осередках щелепних кісток і поділяються на різці, ікла та корінні. Ротовий отвір оточений м'ясистими губами, що властиво лише ссавцям у зв'язку з вигодовуванням молоком. У ротовій порожнині їжа, крім прожовування зубами, піддається хімічному впливу ферментів слини, а потім послідовно переходить у стравохід та шлунок. Шлунок у ссавців добре відокремлений від інших відділів травного тракту та забезпечений травними залозами. У більшості видів ссавців шлунок розділений на більшу чи меншу кількість відділів. Найбільш складний він у жуйних парнокопитних. Кишечник має тонкий та товстий відділи. На межі тонкого та товстого відділів відходить сліпа кишка, в якій відбувається зброджування клітковини. Протоки печінки та підшлункової залози відкриваються в порожнину дванадцятипалої кишки.
Запитання 28
Ендокринна система.
У будь-якому організмі виробляються сполуки, що розносяться по всьому організму, що мають інтегративну роль. У рослин є фітогормони, що контролюють зростання, розвиток плодів, квітів, розвиток пазушних бруньок, розподіл камбію та ін. Фітогормони є у одноклітинних водоростей.
Гормони з'явилися у багатоклітинних організмів, коли з'явилися спеціальні ендокринні клітини. Однак хімічні сполуки, які відіграють роль гормонів, були й раніше. Тироксин, трийодтиронін (щитовидна залоза) виявлено у ціанобактерій. Гормональна регуляція у комах вивчена погано.
В 1965 Вільсон виділив інсулін з морської зірки.
Виявилося, що визначення гормону дуже важко.
Гормон- це специфічна хімічна речовина, що виділяється особливими клітинами в певній ділянці тіла, яка надходить у кров і потім специфічно впливає на певні клітини або органи-мішені, розташовані в інших областях тіла, що призводить до координації функцій всього організму в цілому.
Відома велика кількість гормонів ссавців. Вони поділяються на 3 основні групи.
Феромони. Виділяються у зовнішнє середовище. З їх допомогою тварини приймають та передають інформацію. У людини запах 14 – окситететрадеканової кислоти чітко розрізняють лише жінки, які досягли статевої зрілості.
Найбільш просто організовані багатоклітинні організми – наприклад, губки теж мають подібність до ендокринної системи. Губки складаються з 2 шарів – ентодермою та екзодермою, між ними розташовується мезенхіма, в якій містяться макромолекулярні сполуки, характерні для сполучної тканини високоорганізованих організмів. У мезенхімі є мігруючі клітини, деякі клітини здатні секретувати серотонію, ацетилхолін. Нервова система у губок відсутня. Речовини, синтезовані в мезенхімі, служать зв'язку окремих елементів організму. Координація здійснюється за рахунок переміщення клітин по мезенхімі. Є також перенесення речовин між клітинами. Закладено основу хімічної сигналізації, що у інших тварин. Самостійних ендокринних клітин немає.
У кишковопорожнинних є примітивна нервова система. Спочатку нервові клітини виконували нейросекреторну функцію. Трофічну функцію здійснювали контроль зростання, розвитку організму. Потім нервові клітини стали витягуватись і утворили довгі відростки. Секрет виділявся у органа-мішені, без перенесення (т.к. був крові). Ендокринний механізм виник раніше провідникового. Нервові клітини були ендокринними, а потім отримали провідникові властивості. Нейросекреторні клітини були першими секреторними клітинами.
Первиннороті та вториннороті виробляють однакові стероїдні та пептидні гормони. Вважають, що у процесі еволюції з одних полипептидных гормонів можуть виникнути нові (мутації, дуплікації генів). Дуплікації менш пригнічуються природним відбором, ніж мутації. Багато гормонів можу синтезуватися над одній залозі, а кількох. Наприклад, інсулін виробляється в підшлунковій залозі, підщелепній залозі, 12-палій кишці та інших органах. Існує залежність генів, які контролюють синтез гормонів від становища.
