Regenerarea ca proprietate a viețuitoarelor: capacitatea de auto-reînnoire și restaurare. Tipuri de regenerare
REGENERARE
restaurarea de către organism a părților pierdute la una sau alta etapă a ciclului de viață. Regenerarea are loc de obicei în caz de deteriorare sau pierdere a unui organ sau a unei părți a corpului. Cu toate acestea, pe lângă aceasta, procesele de restaurare și reînnoire au loc în mod constant în fiecare organism de-a lungul vieții sale. La oameni, de exemplu, stratul exterior al pielii este reînnoit constant. Păsările își aruncă periodic pene și cresc altele noi, iar mamiferele își schimbă blana. Copacii de foioase își pierd frunzele în fiecare an și sunt înlocuiți cu altele proaspete. O astfel de regenerare, care de obicei nu este asociată cu daune sau pierderi, se numește fiziologică. Regenerarea care apare după deteriorarea sau pierderea oricărei părți a corpului se numește reparatorie. Aici vom lua în considerare doar regenerarea reparatorie. Regenerarea reparatorie poate fi tipica sau atipica. În regenerarea tipică, partea pierdută este înlocuită cu dezvoltarea exact aceleiași părți. Cauza pierderii poate fi o forță externă (de exemplu, amputarea), sau animalul își poate rupe în mod deliberat o parte a corpului (autotomie), precum o șopârlă care își rupe o parte din coadă pentru a scăpa de un inamic. Cu regenerarea atipică, partea pierdută este înlocuită cu o structură care diferă de originală cantitativ sau calitativ. Membrul regenerat al unui mormoloc poate avea mai puține degete de la picioare decât cel original, iar unui creveți poate crește o antenă în loc de un ochi amputat.
REGENERAREA LA ANIMALE
Capacitatea de regenerare este larg răspândită printre animale. În general, animalele inferioare sunt mai des capabile de regenerare decât formele mai complexe, foarte organizate. Astfel, printre nevertebrate există mult mai multe specii capabile să restabilească organele pierdute decât printre vertebrate, dar numai la unele dintre ele este posibilă regenerarea unui întreg individ dintr-un mic fragment. Cu toate acestea, regula generală că capacitatea de regenerare scade odată cu creșterea complexității organismului nu poate fi considerată absolută. Astfel de animale primitive precum ctenoforele și rotiferele sunt practic incapabile de regenerare, dar la crustacee și amfibieni mult mai complexe această capacitate este bine exprimată; Sunt cunoscute și alte excepții. Unele animale strâns înrudite diferă foarte mult în acest sens. Astfel, într-un râme, un nou individ se poate regenera complet dintr-o mică parte a corpului său, în timp ce lipitorile nu sunt capabile să restaureze un organ pierdut. La amfibienii cu coadă, se formează un membru nou în locul membrului amputat, dar la broaște, ciotul pur și simplu se vindecă și nu are loc o nouă creștere. Multe nevertebrate sunt capabile să regenereze părți mari ale corpului lor. În bureți, polipi hidroizi, viermi plati, tenii și anelide, briozoare, echinoderme și tunicate, un întreg organism se poate regenera dintr-un mic fragment de corp. Deosebit de remarcată este capacitatea de a se regenera în bureți. Dacă corpul unui burete adult este presat prin țesutul plasă, atunci toate celulele se vor separa unele de altele, ca și cum ar fi cernute printr-o sită. Dacă apoi puneți toate aceste celule individuale în apă și amestecați cu grijă, bine, distrugând complet toate conexiunile dintre ele, atunci după un timp încep să se apropie treptat și să se reunească, formând un burete întreg, asemănător celui precedent. Aceasta implică un fel de „recunoaștere” la nivel celular, așa cum demonstrează următorul experiment. Bureții din trei specii diferite au fost separați în celule separate în modul descris și amestecați bine. În același timp, s-a descoperit că celulele fiecărei specii sunt capabile să „recunoaște” celulele propriei specii în masa totală și să se reunească numai cu ele, astfel încât, ca urmare, nu unul, ci trei bureți noi au fost. format, asemănător celor trei originale.
Tenia, care este de multe ori mai lungă decât lată, poate recrea un întreg individ din orice parte a corpului său. Teoretic este posibil, prin tăierea unui vierme în 200.000 de bucăți, să se obțină din acesta 200.000 de viermi noi ca urmare a regenerării. Dintr-o rază de stea de mare, o stea întreagă se poate regenera.

Moluștele, artropodele și vertebratele nu sunt capabile să regenereze un individ întreg dintr-un singur fragment, cu toate acestea, în multe dintre ele organul pierdut este restaurat. Unii recurg la autotomie dacă este necesar. Păsările și mamiferele, ca cele mai avansate animale din punct de vedere evolutiv, sunt mai puțin capabile de regenerare decât altele. La păsări, este posibil să înlocuiți penele și unele părți ale ciocului. Mamiferele își pot restaura tegumentul, ghearele și parțial ficatul; sunt, de asemenea, capabili să vindece rănile, iar căprioarele sunt capabile să crească coarne noi pentru a le înlocui.
Procese de regenerare. Două procese sunt implicate în regenerarea animalelor: epimorfoza și morfalaxia. În regenerarea epimorfă, partea pierdută a corpului este restabilită datorită activității celulelor nediferențiate. Aceste celule asemănătoare embrionului se acumulează sub epiderma rănită la suprafața tăiată, unde formează primordiul sau blastema. Celulele blasteme se înmulțesc treptat și se transformă în țesutul unui nou organ sau parte a corpului. În morfalaxie, alte țesuturi ale corpului sau organului sunt direct transformate în structurile părții lipsă. La polipii hidroizi, regenerarea are loc în principal prin morfalaxie, în timp ce la planari atât epimorfoza, cât și morfalaxia sunt implicate simultan în ea. Regenerarea prin formarea blastemului este larg răspândită la nevertebrate și joacă un rol deosebit de important în regenerarea organelor la amfibieni. Există două teorii cu privire la originea celulelor blastem: 1) celulele blastem provin din „celule de rezervă”, adică. celule care au rămas neutilizate în timpul dezvoltării embrionare și au fost distribuite între diferite organe ale corpului; 2) țesuturi, a căror integritate a fost încălcată în timpul amputării, „dediferențiate” în zona inciziei, adică. se dezintegrează și se transformă în celule blastomatice individuale. Astfel, conform teoriei „celulei de rezervă”, blastemul este format din celule care au rămas embrionare, care migrează din diferite părți ale corpului și se acumulează în apropierea suprafeței tăiate, iar conform teoriei „țesutului dediferențiat”, celulele blastemului provin din celulele țesuturilor deteriorate. Există suficiente date pentru a susține atât una, cât și cealaltă teorie. De exemplu, la planari, celulele de rezervă sunt mai sensibile la razele X decât celulele de țesut diferențiat; prin urmare, ele pot fi distruse prin dozarea strictă a radiațiilor pentru a nu deteriora țesutul planar normal. Indivizii iradiați în acest fel supraviețuiesc, dar își pierd capacitatea de regenerare. Cu toate acestea, dacă numai jumătatea anterioară a corpului planar este iradiată și apoi tăiată, atunci are loc regenerarea, deși cu o oarecare întârziere. Întârzierea indică faptul că blastema este format din celulele de rezervă care migrează către suprafața tăiată din jumătatea neiradiată a corpului. Migrarea acestor celule de rezervă în toată partea iradiată a corpului poate fi observată la microscop. Experimente similare au arătat că la triton, regenerarea membrelor are loc datorită celulelor blastem de origine locală, adică. datorită dediferențierii țesuturilor ciotului deteriorate. Dacă, de exemplu, iradiați întreaga larvă de triton, cu excepția, de exemplu, membrului anterior drept, și apoi amputați acel membru la nivelul antebrațului, animalul va crește un nou membru anterior. Este evident că celulele blastema necesare pentru aceasta provin tocmai din ciotul membrului anterior, deoarece restul corpului a fost iradiat. Mai mult, regenerarea are loc chiar daca intreaga larva este iradiata, cu exceptia unei zone late de 1 mm pe tarsul anterior drept, iar apoi acesta din urma este amputat prin efectuarea unei incizii prin aceasta zona neiradiata. În acest caz, este destul de clar că celulele blastemului provin de la suprafața tăiată, deoarece întregul corp, inclusiv piciorul drept, a fost lipsit de capacitatea de a se regenera. Procesele descrise au fost analizate folosind metode moderne. Un microscop electronic vă permite să observați modificările în țesuturile deteriorate și regenerabile în toate detaliile. Au fost creați coloranți care dezvăluie anumite substanțe chimice conținute în celule și țesuturi. Metodele histochimice (folosind coloranți) fac posibilă evaluarea proceselor biochimice care au loc în timpul regenerării organelor și țesuturilor.
Polaritate. Una dintre cele mai misterioase probleme din biologie este originea polarității în organisme. Din oul sferic al unei broaște se dezvoltă un mormoloc, care de la bun început are un cap cu creier, ochi și gură la un capăt al corpului și o coadă la celălalt. În mod similar, dacă tăiați corpul unei planari în fragmente individuale, se dezvoltă un cap la un capăt al fiecărui fragment și o coadă la celălalt. În acest caz, capul este întotdeauna format la capătul anterior al fragmentului. Experimentele arată clar că planaria are un gradient de activitate metabolică (biochimică) de-a lungul axei anterioare-posterior a corpului său; în acest caz, cea mai mare activitate este la extremitatea anterioră a corpului, iar spre capătul posterior activitatea scade treptat. La orice animal, capul se formează întotdeauna la capătul fragmentului unde activitatea metabolică este mai mare. Dacă direcția gradientului activității metabolice într-un fragment izolat de planaria este inversată, atunci formarea capului va avea loc la capătul opus al fragmentului. Gradientul activității metabolice în corpul planariilor reflectă existența unui gradient fizico-chimic mai important, a cărui natură este încă necunoscută. În limbul regenerant al unui triton, polaritatea structurii nou formate pare să fie determinată de ciotul conservat. Din motive care rămân încă neclare, în organul în regenerare se formează doar structuri situate distal de suprafața plăgii, iar cele situate mai proximal (mai aproape de corp) nu se regenerează niciodată. Deci, dacă mâna unui triton este amputată, iar partea rămasă a membrului anterior este introdusă cu capătul tăiat în peretele corpului și acest capăt distal (depărtat de corp) este lăsat să prindă rădăcini într-un loc nou, neobișnuit pentru acesta, apoi transecția ulterioară a acestui membru superior în apropierea umărului (eliberându-l de legătura cu umărul) duce la regenerarea membrului cu un set complet de structuri distale. În momentul tăierii, un astfel de membru are următoarele părți (începând de la încheietura mâinii, fuzionate cu peretele corpului): încheietura mâinii, antebrațul, cotul și jumătatea distală a umărului; apoi, ca urmare a regenerării, apar următoarele: o altă jumătate distală a umărului, cotului, antebrațului, încheieturii mâinii și mâinii. Astfel, membrul inversat (cu susul în jos) a regenerat toate părțile situate distal de suprafața plăgii. Acest fenomen izbitor indică faptul că țesuturile ciotului (în acest caz ciotul membrului) controlează regenerarea organului. Sarcina cercetărilor ulterioare este de a afla exact ce factori controlează acest proces, ce stimulează regenerarea și ce provoacă acumularea celulelor care asigură regenerarea pe suprafața plăgii. Unii oameni de știință cred că țesutul deteriorat eliberează un fel de „factor al plăgii” chimic. Cu toate acestea, încă nu a fost posibilă izolarea unei substanțe chimice specifice rănilor.
REGENERAREA ÎN PLANTE
Apariția pe scară largă a regenerării în regnul vegetal se datorează conservării meristemelor (țesuturilor formate din celule în diviziune) și a țesuturilor nediferențiate. În cele mai multe cazuri, regenerarea la plante este, în esență, una dintre formele de înmulțire vegetativă. Astfel, la varful unei tulpini normale se afla un mugure apical, care asigura formarea continua a frunzelor noi si cresterea tulpinii in lungime pe toata durata de viata a plantei. Dacă acest mugure este tăiat și menținut umed, se dezvoltă adesea rădăcini noi din celulele parenchimului prezente în el sau din calusul format pe suprafața tăieturii; mugurele continuă să crească și dă naștere unei noi plante. Același lucru se întâmplă în natură când o ramură se rupe. Genele și stolonii sunt separați ca urmare a morții secțiunilor vechi (internoduri). În același mod, rizomii de iris, picior de lup sau ferigi sunt împărțiți, formând noi plante. De obicei, tuberculii, cum ar fi tuberculii de cartofi, continuă să trăiască după ce tulpina subterană pe care au crescut a murit; odată cu debutul unui nou sezon de creștere, pot da naștere propriilor rădăcini și lăstari. La plantele cu bulbi, cum ar fi zambilele sau lalelele, la baza solzilor bulbilor se formează lăstari și pot forma, la rândul lor, bulbi noi, care în cele din urmă produc rădăcini și tulpini florale, de exemplu. devin plante independente. La unii crini se formează bulbi aerieni la axilele frunzelor, iar la o serie de ferigi, pe frunze cresc muguri de puiet; la un moment dat cad la pământ și își reiau creșterea. Rădăcinile sunt mai puțin capabile să formeze părți noi decât tulpinile. Pentru aceasta, tuberculul de dalie are nevoie de un mugure care se formează la baza tulpinii; cu toate acestea, cartofii dulci pot da naștere unei noi plante dintr-un mugure format dintr-un con de rădăcină. Frunzele sunt, de asemenea, capabile de regenerare. La unele specii de ferigi, de exemplu, la feriga (Camptosorus), frunzele sunt foarte alungite și arată ca niște structuri asemănătoare părului lung care se termină într-un meristem. Din acest meristem se dezvoltă embrionul cu tulpină, rădăcini și frunze rudimentare; dacă vârful frunzei plantei părinte se îndoaie și atinge solul sau mușchiul, mugurul începe să crească. Noua plantă se separă de părinte după epuizarea acestei formațiuni asemănătoare părului. Frunzele plantei de apartament suculente Kalanchoe poartă pe margini plantule bine dezvoltate care cad ușor. Pe suprafața frunzelor de begonie se formează lăstari și rădăcini noi. Pe frunzele unor mușchi de club (Lycopodium) și hepatice (Marchantia) se dezvoltă corpuri speciale numite muguri embrionari; căzând la pământ, prind rădăcini și formează noi plante mature. Multe alge se reproduc cu succes prin spargerea în fragmente sub impactul valurilor.
Vezi si SISTEMATICA PLANTELOR. LITERATURA Mattson P. Regenerare - prezent și viitor. M., 1982 Gilbert S. Developmental biology, voi. 1-3. M., 1993-1995
Enciclopedia lui Collier. - Societate deschisă. 2000 .
Sinonime:Vedeți ce este „REGENERARE” în alte dicționare:
REGENERARE- REGENERARE, procesul de formare a unui nou organ sau țesut în locul unei părți a corpului care a fost îndepărtată într-un fel sau altul. De foarte multe ori R. este definit ca fiind procesul de refacere a ceea ce s-a pierdut, adică formarea unui organ asemănător celui îndepărtat. Acest... ... Marea Enciclopedie Medicală
- (lat. târzie, din lat. re din nou, din nou, și gen, eris gen, generație). Reînnoirea, reînnoirea, restaurarea a ceea ce a fost distrus. În sens figurat: o schimbare în bine. Dicționar de cuvinte străine incluse în limba rusă.... ... Dicționar de cuvinte străine ale limbii ruse
REGENERARE, în biologie, capacitatea organismului de a înlocui una dintre părțile pierdute. Termenul de regenerare se referă și la o formă de reproducere asexuată în care un nou individ ia naștere dintr-o parte separată a corpului mamei... Dicționar enciclopedic științific și tehnic
Restaurare, recuperare; compensare, regenerare, reînnoire, heteromorfoză, pettencoferare, renaștere, morfalaxie Dicționar de sinonime rusești. regenerare substantiv, număr de sinonime: 11 compensare (20) ... Dicţionar de sinonime
1) refacerea, prin anumite procedee fizico-chimice, a compoziției și proprietăților originale ale deșeurilor pentru reutilizarea acestora. În afacerile militare, regenerarea aerului a devenit larg răspândită (în special pe subacvatice... ... Dicționar marin
Regenerare- – readucerea produsului folosit la proprietățile sale originale. [Dicționar terminologic de beton și beton armat. FSUE „Centrul de cercetare „Construcții” NIIZHB numit după. A. A. Gvozdeva, Moscova, 2007, 110 p.] Regenerare - restaurarea deșeurilor... ... Enciclopedie de termeni, definiții și explicații ale materialelor de construcție
REGENERARE- (1) refacerea proprietăților și compoziției originale a deșeurilor (apă, aer, uleiuri, cauciuc etc.) pentru reutilizarea acestora. Se realizează cu ajutorul anumitor fizice chimic. procese în dispozitive speciale de regenerare. Lat...... Marea Enciclopedie Politehnică
- (din lat. târzie regeneratio rebirth renewal), în biologie, restaurarea de către organism a organelor și țesuturilor pierdute sau deteriorate, precum și refacerea întregului organism din partea sa. Caracteristic în principal plantelor și nevertebratelor... ...
În tehnologie, 1) readucerea produsului uzat la calitățile sale originale, de exemplu. restaurarea proprietăților nisipului uzat de turnare în turnătorii, purificarea uleiului lubrifiant uzat, transformarea produselor din cauciuc uzate în plastic... ... Dicţionar enciclopedic mare
REGENERARE, regenerare, multe. nu, femeie (lat. regeneratio restaurare, retur). 1. Încălzirea gazului și a aerului care intră în cuptor cu produse de ardere reziduale (tehnice). 2. Reproducerea organelor pierdute de către animale (zool.). 3. Radiația...... Dicționarul explicativ al lui Ushakov
Regenerare(din lat. regeneratio- renașterea) este procesul de refacere a structurilor biologice în timpul vieții unui organism. Regenerarea menține structura și funcțiile organismului, integritatea acestuia.Procesele de regenerare sunt implementate la diferite niveluri de organizare - genetică moleculară, subcelulară, celulară, tisulară, de organ, organism.La nivel genetic molecular, replicarea ADN-ului, repararea acestuia, sinteza de se efectuează noi enzime, molecule de ATP etc. Toate aceste procese sunt incluse în metabolismul celulei.La nivel subcelular, structurile celulare sunt restaurate datorită formării de noi unități structurale și a ansamblării organitelor sau a divizării organitelor supraviețuitoare. De exemplu, structurile mobile ale membranei celulare - receptori, canale ionice și pompe - se pot mișca, concentra sau distribui în interiorul membranei. În plus, părăsesc membrana, sunt distruse și înlocuite cu altele noi. Astfel, în mioblaste, aproximativ 1 µm2 din suprafață este degradat și înlocuit cu noi molecule în fiecare minut. În celulele fotoreceptoare - tije (Fig. 8.73) există un segment exterior format din aproximativ o mie de așa-numitele discuri fotoreceptoare - secțiuni dens împachetate ale membranei celulare în care sunt scufundate proteinele sensibile la lumină asociate pigmentului vizual. Aceste discuri sunt reînnoite continuu - se degradează la capătul exterior și reapar la capătul interior cu o viteză de 3-4 discuri pe oră. Procesele de recuperare după deteriorare se desfășoară într-un mod similar. Expunerea la otrăvuri mitocondriale determină pierderea crestaților mitocondriale. Dupa incetarea actiunii otravii in celula hepatica, mitocondriile isi refac structura in 2-3 zile.Nivelul celular de regenerare presupune refacerea structurii si, in unele cazuri, a functiilor celulei. Exemple de acest fel includ refacerea procesului unei celule nervoase a unui neuron. La mamifere, acest proces are loc cu o rată de 1 mm pe zi. Restaurarea funcțiilor celulare poate fi realizată prin hiperplazie- cresterea numarului de organele intracelulare (regenerare intracelulara).La urmatorul nivel - tesutul sau populatia celulara - are loc refacerea celulelor pierdute de o anumita directie de diferentiere. Restructurarile apar in cadrul populatiilor de celule, iar rezultatul lor este refacerea functiilor tisulare. Astfel, la om, durata de viață a celulelor epiteliale intestinale este de 4-5 zile, trombocitele - 5-7 zile, eritrocitele - 120-125 zile. În fiecare secundă, aproximativ 1 milion de globule roșii sunt distruse și aceeași cantitate se formează din nou în măduva osoasă roșie. Capacitatea de a reface celulele pierdute este asigurată datorită faptului că există două compartimente celulare în țesuturi. Una este celulele lucrătoare diferențiate, iar cealaltă este celulele cambiale capabile de divizare și diferențiere ulterioară. Acestea din urmă sunt numite în prezent celule stem regionale (vezi paragrafele 3.1.2, 3.2). Ei sunt comisi, i.e. soarta lor este predeterminată (a se vedea secțiunea 8.3.1), prin urmare sunt capabile să dea naștere la unul sau mai multe tipuri de celule specifice. Diferențierea lor ulterioară este determinată de semnale care vin din exterior: din mediu (interacțiuni intercelulare) și semnale la distanță (de exemplu, hormoni), în funcție de genele specifice care sunt activate selectiv în celule. Astfel, în epiteliul intestinului subțire, celulele cambiale sunt situate în zonele inferioare ale criptelor (Fig. 8.74). Sub anumite influențe, ele sunt capabile să dea naștere la celule ale epiteliului de absorbție „marginal” și a unor glande unicelulare Nivelul de regenerare a organului implică restabilirea funcției sau structurii organului. La acest nivel se observă nu numai transformări ale populațiilor celulare, ci și procese morfogenetice. În acest caz, sunt implementate aceleași mecanisme ca și în timpul formării organelor în embriogeneză. Ta- Orez. 8,73. Reprezentarea schematică a fotoreceptorului retinian - tije: 1 - corp sinaptic adiacent stratului neural al retinei, 2 - nucleu, 3 - aparatul Golgi, 4 - segment interior cu mitocondrii, 5 - cili de legătură, 6 - segment exterior cu discuri fotoreceptoare prin ce fel de regenerare poate fi efectuatăepimorfoză, morfolaxie, hipertrofie regenerativă.Acestemetodele și mecanismele de regenerare sunt discutate mai jos. La nivel de organism, în unele cazuri este posibil să se recreeze un întreg organism dintr-una sau un grup de celule. Există două tipuri de regenerare:fiziologicȘireparatoriu.Regenerare fiziologică (homeostatică). este procesul de restaurare a structurilor care se uzează în timpul vieții normale. Datorită acesteia, homeostazia structurală este menținută și organele își pot îndeplini în mod constant funcțiile. Din punct de vedere biologic general, regenerarea fiziologică, ca și metabolismul, este o manifestare a unei proprietăți atât de importante a vieții precum auto-înnoirea. Auto-reînnoirea asigură existența organismului în timp și spațiu. Se bazează pe migrarea biogenă a atomilor. La nivel intracelular, importanța regenerării fiziologice este deosebit de mare pentru așa-numitele țesuturi „eterne” care și-au pierdut capacitatea de a se regenera prin diviziunea celulară. În primul rând, acest lucru se aplică țesutului nervos, retinei. La nivel celular și tisular, regenerarea fiziologică are loc în țesuturile „labile”, unde Orez. 8,74. Localizarea celulelor stem regionale în epiteliul intestinului subțire: 1 - celule nedivizate; 2 - divizarea celulelor stem; 3 - celule cu diviziune rapidă; 4 - celule diferențiate nedivizate; 5 — direcția mișcării celulei; 6 - celule exfoliate de pe suprafața vilozităților intestinale; intensitatea reînnoirii celulare este foarte mare, iar în țesuturile „în creștere”, ale căror celule se reînnoiesc mult mai lent. Primul grup include, de exemplu, corneea ochiului, epiteliul mucoasei intestinale, celulele sanguine periferice, epiderma pielii și derivații săi - păr și unghii. Celulele organelor precum ficatul, rinichii și glanda suprarenală constituie al doilea dintre aceste grupuri.Intensitatea proliferării este judecată după numărul de mitoze la 1000 de celule numărate. Dacă luăm în considerare că mitoza în sine durează în medie aproximativ 1 oră, iar întregul ciclu mitotic din celulele somatice durează în medie 22-24 de ore, devine clar că pentru a determina intensitatea reînnoirii compoziției celulare a țesuturilor, este necesară numărarea numărului de mitoze pe parcursul uneia sau mai multor zile. S-a dovedit că numărul de celule care se divid nu este același în diferite momente ale zilei. Așa a fost descoperit ritmul zilnic al diviziunii celulare, un exemplu al căruia este prezentat în Fig. 8.75.Un ritm zilnic al numărului de mitoze a fost găsit nu numai în țesuturile normale, ci și în țesuturile tumorale. Reflectă un model mai general, Orez. 8,75. Modificări zilnice ale indicelui mitotic (MI) în epiteliul esofagului (1) și corneei (2) la șoareci. Indicele mitotic este exprimat în ppm (0/00), reflectând numărul de mitoze dintr-o mie de celule numărate și anume ritmul tuturor funcțiilor corpului. Una dintre domeniile moderne ale biologiei estecronobiologie— studiază, în special, mecanismele de reglare a ritmurilor zilnice ale activității mitotice, care este de mare importanță pentru medicină. Existența unei periodicități zilnice a numărului de mitoze indică ajustabilitatea regenerării fiziologice de către organism. Pe lângă ciclurile zilnice, există cicluri lunare și anuale de reînnoire a țesuturilor și a organelor. Regenerarea fiziologică este inerentă organismelor tuturor speciilor, dar are loc mai ales intens la vertebratele cu sânge cald, deoarece acestea au, în general, o intensitate foarte mare de funcționare a tuturor organelor în comparație cu alte animale. Regenerare reparatorie(din lat.reparatio - restaurare) - refacerea structurilor biologice după leziuni și alți factori vătămatori. Astfel de factori pot include substanțe toxice, agenți patogeni, temperaturi ridicate și scăzute (arsuri și degerături), expunerea la radiații, postul etc. Capacitatea de regenerare nu are o dependență clară de nivelul de organizare, deși s-a remarcat de mult timp că animalele organizate inferioară au o capacitate mai bună de a regenera organele externe. Acest lucru este confirmat de exemple uimitoare de regenerare a hidrelor, planarelor, anelidelor, artropodelor, echinodermelor și cordatelor inferioare, cum ar fi ascidie. Dintre vertebrate, amfibienii cu coadă au cea mai bună capacitate de regenerare. Se știe că diferite specii din aceeași clasă pot diferi foarte mult în ceea ce privește capacitatea lor de a se regenera. În plus, la studierea capacității de a regenera organele interne, s-a dovedit că este semnificativ mai mare la animalele cu sânge cald, cum ar fi mamiferele, în comparație cu amfibieni. Regenerarea la mamifere este unică. Pentru regenerarea unor organe externe sunt necesare condiții speciale. Limba și urechea, de exemplu, nu se regenerează cu leziuni marginale (de fapt, vorbim despre amputarea părții marginale a structurii). Dacă aplicați un defect traversant pe toată grosimea organului, recuperarea merge bine. Regenerarea organelor interne poate fi foarte activă. Un întreg organ este restaurat dintr-un mic fragment de ovar. Există o presupunere că imposibilitatea regenerării membrelor și a altor organe externe la mamifere este de natură adaptativă și se datorează selecției, deoarece cu un stil de viață activ, procesele morfogenetice care necesită o reglare complexă ar îngreuna existența. O serie de cercetători cred că organismele aveau inițial două moduri de vindecare a rănilor - acțiunea sistemului imunitar și regenerare. Dar în cursul evoluției au devenit incompatibile între ele. În timp ce regenerarea poate părea o alegere mai bună, ceea ce este mai important pentru noi sunt celulele T ale sistemului imunitar - arma noastră principală împotriva tumorilor. Regenerarea unui membru devine lipsită de sens dacă celulele canceroase se dezvoltă rapid în organism în același timp. Se pare că sistemul imunitar, în timp ce ne protejează de infecții și cancer, suprimă simultan capacitatea noastră de recuperare.Volumul de regenerare reparatorie poate fi foarte diferit.O opțiune extremă este restaurarea întregului organism dintr-o mică parte separată a acestuia. , de fapt dintr-un grup de celule somatice. Printre animale, o astfel de restaurare este posibilă în bureți și celenterate. Hidra poate fi regenerată dintr-un grup de celule obținute prin presarea ei printr-o sită. Dintre plante, dezvoltarea unei plante cu totul noi este posibilă chiar și dintr-o celulă somatică, așa cum s-a obținut cu exemplul morcovilor și tutunului. Acest tip de procese de restaurare este însoțit de apariția unei noi axe morfogenetice a corpului și poartă denumirea de B.P. Tokin „embriogeneză somatică”, deoarece în multe privințe seamănă cu dezvoltarea embrionară. Ca o opțiune similară pentru regenerare, poate fi luată în considerare clonarea experimentală a unui întreg organism dintr-o celulă somatică la mamifere.Următoarea opțiune în ceea ce privește domeniul de aplicare este restaurarea unor zone mari ale corpului, constând dintr-un complex de organe. Un exemplu este regenerarea în hidre, vierme ciliat (planaria) și stele de mare (Fig. 8.76). Îndepărtând o parte a animalului din fragmentul rămas, chiar și unul foarte mic, este posibilă restabilirea unui organism cu drepturi depline. De exemplu, restaurarea unei stele de mare dintr-o rază conservată Următoarea în această serie este regenerarea organelor individuale, care este larg răspândită în regnul animal, de exemplu, coada unei șopârle, ochii artropodelor, ochiul, membrele , și coada unui triton Vindecarea pielii, rănilor, rănilor oaselor și a altor organe interne - cel mai puțin amplu proces, dar nu mai puțin important pentru restabilirea integrității structurale și funcționale a corpului Există mai multe metode de regenerare reparatorie. Acestea includ epimorfoza, morfalaxia, hipertrofia regenerativă, hipertrofia compensatorie, vindecarea rănilor epiteliale, regenerarea tisulară. Orez. 8,76. Regenerarea unui complex de organe la unele specii de animale nevertebrate: a - hidra; b - vierme plat; c - stea de mare; d - restaurarea unei stele de mare dintr-o rază Epimorfoza este cea mai evidentă metodă de regenerare, constând în creșterea unui nou organ de pe suprafața amputației. O ilustrare este regenerarea cristalinului sau a membrelor la amfibienii cu coadă (Fig. 8.77). Să luăm în considerare mai detaliat procesul de regenerare folosind exemplul de epimorfoză a membrului tritonului. În timpul procesului de recuperare se disting fazele regresive și progresive de regenerare. Faza de regresie începe cu vindecarea rănilor, timp în care apar următoarele evenimente principale: oprire Orez. 8,77. Regenerarea cristalinului (1) din irisul dorsal (2) în sângerare triton, contracția țesuturilor moi ale ciotului membrului, formarea unui cheag de fibrină pe suprafața plăgii și migrarea epidermei care acoperă suprafața amputației. Apoi distrugerea țesutului începe imediat proximal de locul amputației. În același timp, celulele implicate în procesul inflamator pătrund în țesuturile moi distruse, se observă fagocitoză și edem local. În urma acesteia, începe dediferențierea celulelor specializate în zona de sub epiderma plăgii: mușchi, os, cartilaj etc. Celulele capătă caracteristici mezenchimale, formează un grup și se formează blastem regenerativ(Fig. 8.78). În același timp, epiderma plăgii se îngroașă și se formează rapid capac ectodermic apical.În această etapă, vasele și fibrele nervoase cresc în blastoma regenerativă și calota ectodermică. În continuare, începe faza progresivă, care se caracterizează cel mai mult prin procesele de creștere și morfogeneză. Lungimea și greutatea blastemului regenerativ crește rapid. Ia o formă conică. Celulele mezenchimale ale blastemului se dediferențiază, dând naștere tuturor tipurilor de celule specializate care sunt necesare pentru a forma structurile membrului. Au loc creșterea membrului și morfogeneza acestuia (formarea formei). Când forma generală a membrului s-a dezvoltat deja, regenerarea este încă mai mică decât membrul normal. Cu cât animalul este mai mare, cu atât este mai mare această diferență de mărime. Finalizarea morfogenezei necesită timp, după care regenerarea ajunge la dimensiunea unui membru normal.În Fig. 8,79. Orez. 8,78. Regenerarea unui membru la un triton: a - membru normal, b - amputație; c — formarea calotei apicale și a blastemului; d — rediferențierea celulelor; d - membru nou format. 1 - blastemă; 2 - calota ectodermica apicala; 3 - rediferențierea celulelor blastematice (explicații în text) La larvele tinere de axolotl, limbul se poate regenera în 3 săptămâni, la tritoni și axoloți adulți - în 1-2 luni, iar la ambistos terestru acest lucru durează aproximativ 1 an. Morfalaxie— regenerare prin restructurarea zonei de regenerare. Un exemplu este regenerarea unei hidre dintr-un inel tăiat din mijlocul corpului sau restaurarea unei planarii dintr-o zecime sau douăzecime din partea sa. În acest caz, nu au loc procese semnificative de modelare pe suprafața plăgii. Piesa tăiată se micșorează, celulele din interiorul ei se rearanjează și apare un întreg individ de dimensiuni reduse, care apoi crește. Această metodă de regenerare a fost descrisă pentru prima dată de T. Morgan în 1900. În conformitate cu descrierea sa, morfalaxia are loc fără mitoză. Adesea există o combinație de creștere epimorfă la locul amputației cu reorganizare prin morfalaxie în părțile adiacente ale corpului. Hipertrofie regenerativă (endomorfoză) se referă la organele interne. Această metodă de regenerare presupune creșterea dimensiunii organului rămas fără a-i restabili forma inițială. O ilustrare este regenerarea ficatului vertebratelor, inclusiv mamiferelor. Cu o leziune marginală a ficatului, partea îndepărtată a organului nu este niciodată restaurată. Suprafața rănii se vindecă. În același timp, intern Orez. 8,79. Regenerarea membrului anterior la un triton într-un experiment Orez. 8,80. Efectul vârstei asupra creșterii numărului de glomeruli de nefron după îndepărtarea unui rinichi la șobolani la scurt timp după naștere: 1 - curba creșterii numărului de glomeruli în dezvoltarea normală postnatală la un rinichi; 2 - curbe ale creșterii numărului de glomeruli nou formați după îndepărtarea unui rinichi în diferite stadii de ontogeneză; în partea rămasă, proliferarea celulară (hiperplazie) crește și chiar și după îndepărtarea a 2/3 din ficat, greutatea inițială iar volumul, dar nu forma, sunt restaurate. Structura internă a ficatului se dovedește a fi normală, lobulii au o dimensiune tipică. Funcția ficatului revine, de asemenea, la normal. Hipertrofie compensatorie (vicaria). constă în modificări ale unuia dintre organe cu o încălcare în altul, aparținând aceluiași sistem de organe. Un exemplu este hipertrofia unuia dintre rinichi atunci când celălalt este îndepărtat sau mărirea ganglionilor limfatici când splina este îndepărtată. Modificările capacității pentru acest tip de regenerare în funcție de vârstă sunt prezentate în Fig. 8.80.Ultimele două metode diferă în locul regenerării, dar mecanismele lor sunt aceleaşi: hiperplazie şi hipertrofie (Fig. 8.81)1. 1 Hipertrofie(greacă hiper-+ trofeu— alimentatie, nutritie)- o creștere a volumului și greutății unui organ al corpului sau a părții sale individuale. Hiperplazie (greacă) hiper-+ plasis- formare, formare) - o creștere a numărului de elemente structurale ale țesuturilor prin formarea lor excesivă nouă. Aceasta nu este doar reproducerea celulară, ci și o creștere a ultrastructurilor citoplasmatice (în primul rând mitocondriile, miofilamentele, reticulul endoplasmatic, modificarea ribozomilor). Orez. 8,81. Diagrama care ilustrează mecanismele hipertrofiei și hiperplaziei: a - normal; b - hiperplazie; c — hipertrofie; d - schimbare combinată Epitelizare La vindecarea rănilor cu acoperire epitelială deteriorată, procesul este aproximativ același, indiferent dacă regenerarea organelor are loc în continuare prin epimorfoză sau nu. Vindecarea rănilor epidermice la mamifere, când suprafața plăgii se usucă formând o crustă, se desfășoară după cum urmează (Fig. 8.82). Epiteliul de la marginea plăgii se îngroașă datorită creșterii volumului celular și extinderii spațiilor intercelulare. Cheagul de fibrină joacă rolul de substrat pentru migrarea epidermei în profunzimea plăgii. Nu există mitoze doar în celulele epiteliale migratoare Orez. 8,82. Schema unor evenimente care apar în timpul epitelizării unei plăgi cutanate la mamifere: a - începutul creșterii în interior a epidermei sub țesutul necrotic, b - fuziunea epidermei și separarea crustei; 1 - țesut conjunctiv; 2 - epiderma; 3 - crusta; 4 - țesut necrotic; au activitate fagocitară. Celulele de la marginile opuse vin în contact. Apoi urmează cheratinizarea epidermei plăgii și separarea crustei care acoperă rana. În momentul în care epiderma marginilor opuse se întâlnește în celulele situate imediat în jurul marginii rănii, se observă un focar de mitoze, care apoi dispare treptat.Refacerea țesuturilor mezodermice individuale, cum ar fi musculare și scheletice, se numește regenerarea tesuturilor. Pentru regenerarea musculară este important să se păstreze cel puțin cioturi mici la ambele capete, iar pentru regenerarea osoasă este necesar periostul.Astfel, există multe metode sau tipuri diferite de fenomene morfogenetice în refacerea părților pierdute și deteriorate ale corpului. Diferențele dintre ele nu sunt întotdeauna evidente și este necesară o înțelegere mai profundă a acestor procese.Regenerarea nu produce întotdeauna o copie exactă a structurii îndepărtate. Când tipic regenerare, partea pierdută a structurii corecte este restaurată (homomorfoza), ce nu se întâmplă când atipic regenerare. Un exemplu al acesteia din urmă este apariția unei alte structuri în locul celei pierdute - heteromorfoză. Poate apărea sub formă homeotic regenerare, care constă în apariția unei antene sau a unui membru în locul ochiului la artropode. Altă opțiune - hipomorfoza, regenerare cu înlocuire parțială a structurii amputate. De exemplu, o șopârlă dezvoltă o structură asemănătoare pungii în loc de un membru (Fig. 8.83). Regenerarea atipică poate include cazuri modificări de polaritate structurilor. Astfel, dintr-un fragment scurt de planaria, se poate obține în mod fiabil o planaria bipolară. Are loc formarea de structuri suplimentare sau regenerarea excesivă. După o incizie în bont în timpul amputării secțiunii capului planariei, are loc regenerarea a două sau mai multe capete (Fig. 8.84).Studiul regenerării se referă nu numai la manifestările externe. Există o serie de aspecte care sunt de natură problematică și teoretică. Acestea includ probleme de reglare și condiții în care au loc procesele de restaurare, problemele de origine a celulelor implicate în regenerare, capacitatea de regenerare la diferite grupuri de animale și caracteristicile proceselor de restaurare la mamifere.S-a stabilit că în timpul proceselor de regenerare precum ca determinare, diferențiere și diferențiere, creștere, morfogene- Orez. 8,83. Exemple de regenerare atipică: a - cap normal de cancer; b - formarea unei antene în loc de ochi; c - formarea unei structuri în formă de pungă în locul unui membru într-o salamandră. 1 - ochi; 2 - antena; 3 - locul de amputare; 4 - ganglionul nervos Orez. 8,84. Exemple de regenerare atipică: a - planaria bipolară; b - planaria multicapitala obtinuta dupa amputarea capului si incizii pe culturi, similare proceselor care au loc in dezvoltarea embrionara. Datele obținute până în prezent indică faptul că restaurarea structurilor pierdute se realizează, în esență, pe baza acelorași programe de dezvoltare, care ghidează formarea lor în embrion, și pe baza mecanismelor de dezvoltare celulară și sistemică. Cu toate acestea, în timpul regenerării, toate procesele de dezvoltare apar din nou, adică. într-un organism format, prin urmare restaurarea structurilor are o serie de diferențe și caracteristici specifice. Nu există nicio îndoială că, în timpul regenerării, mecanismele sistemice — interacțiuni intercelulare și interbud, reglarea nervoasă și umorală — joacă o importanță deosebită. Astfel, în timpul epimorfozei membrului tritonului, epiderma formată în timpul epitelizării stimulează liza țesuturilor mezodermice subiacente. În absența acesteia sau când se formează o cicatrice, regenerarea nu are loc. Celulele de sub epiderma formată se dediferențiază și formează un blastem. În această etapă se observă influenţe inductive reciproce între epidermă, care formează capacul ectodermic apical, şi blasmemul mezodermic. În timpul dezvoltării embrionare, în timpul formării membrului, au avut loc interacțiuni similare între mugurele membrului mezodermic și creasta ectodermală apicală. În timpul dediferențierii în celule, activitatea genelor specifice tipului care determină specializarea celulei este suprimată, de exemplu genele MRFȘiMif5în fibrele musculare. Genele necesare pentru proliferarea celulară sunt apoi activate. Unul din eimsx1.În această etapă, procesele nervoase și epiderma care cresc în blastem produc factori trofici și de creștere necesari pentru proliferarea și supraviețuirea celulelor blastemului. Printre acestea se numără factorul de creștere a fibroblastelor FGF-10. Același factor este necesar pentru proliferarea epidermei în sine. Blastem, la rândul său, sintetizează ca răspuns factori neurotrofici care stimulează creșterea nervoasă. Nervii sunt necesari pentru a forma capacul ectodermal apical. În plus, se produce blastoma, ca și capacul epidermic apical FGF-8,care stimulează creșterea capilară în interior. Este de remarcat diferențele observate în această etapă între regenerare și dezvoltarea embrionară. Pentru a implementa regenerarea, este necesară inervația. Fără aceasta, poate apărea dediferențierea celulelor, dar dezvoltarea ulterioară este absentă. În perioada de morfogeneză embrionară a membrului (în timpul diferențierii celulare), nervii nu sunt încă formați. Pe lângă inervație, acțiunea enzimelor metaloproteinaze este necesară în stadiul incipient al regenerării. Ele distrug componentele matricei, ceea ce permite celulelor să se dividă (se disocieze) și să prolifereze activ. Celulele aflate in contact unele cu altele nu pot continua regenerarea si raspund la actiunea factorilor de crestere. Astfel, în timpul regenerării, se observă toate variantele de interacțiuni intercelulare: prin eliberarea de factori paracrini care se difuzează de la o celulă la alta, interacțiunea prin matrice și prin contactul direct al suprafețelor celulare. În timpul etapei de dediferențiere, genele homeotice sunt exprimate în celulele ciotHoxD8ȘiHoxDlO,iar odată cu începutul diferenţierii – geneleHoxD9ȘiHoxD13.După cum sa arătat în secțiunea 8.3.4, aceleași gene sunt transcrise activ în morfogeneza embrionară a membrului. Este important de menționat că în timpul regenerării, diferențierea celulară se pierde, dar determinarea lor este păstrată. Deja în stadiul de blastem nediferențiat, sunt stabilite principalele caracteristici ale membrului regenerant. Acest lucru nu necesită activarea genelor care asigură specificarea membrelor (Tbx-5pentru fata siTbx-4 pentru spate). Membrul se formează în funcție de localizarea blastemului. Dezvoltarea lui se produce la fel ca și în embriogeneză: mai întâi secțiunile proximale, apoi cele distale. Gradientul proximal-distal care determină ce părți ale mugurelui în creștere va deveni un umăr, care va deveni un antebraț și care va deveni o mână este stabilit de gradientul proteic Produsul 1. Este localizat pe suprafața celulelor blastem și concentrația sa este mai mare la baza membrului. Această proteină joacă rolul unui receptor, iar molecula semnal (ligand) pentru aceasta este o proteină ponei. Este sintetizat de celulele Schwann din jurul nervului regenerant. În absența acestei proteine, care prin interacțiunea ligand-receptor declanșează activarea cascadei de gene necesare dezvoltării, regenerarea nu are loc. Acest lucru explică fenomenul de lipsă de restaurare a membrelor atunci când un nerv este tăiat, precum și atunci când un număr insuficient de fibre nervoase crește în blastem. Interesant este că dacă nervul membrului unui triton este retras sub pielea bazei membrului, se formează un membru suplimentar. Dacă este dus la baza cozii, este stimulată formarea unei cozi suplimentare. Reducerea nervului în regiunea laterală nu provoacă structuri suplimentare. Toate acestea au dus la crearea conceptului câmpuri de regenerare. Orez. 8,85. Experimentul cu rotația blastemului membrului (explicații în text) Similar procesului de embriogeneză, axa anterior-posterior se formează și în câmpul membrului în curs de dezvoltare. În rudimentul în curs de dezvoltare apare o zonă de activitate polarizantă, care determină asimetria membrului. Prin rotirea capătului ciotului membrului cu 180°, se poate obține un membru cu dublarea în oglindă a degetelor (Fig. 8.85).Astfel, este adevărat că formarea membrului are loc în câmpul organelor, iar blastemul este un sistem de autoreglare. Alături de cele de mai sus, o dovadă în acest sens este oferită de rezultatele obținute într-o serie de experimente privind transplantul blastemului membrului anterior pe blastemul de la mijlocul coapsei (Fig. 8.86). Atunci când este transplantată în câmpul de regenerare al altui membru, grefa este poziționată în conformitate cu informațiile de poziție primite (gradienți de substanță): blastoma umărului este deplasată la mijlocul coapsei, antebrațul - spre partea inferioară a piciorului, încheietura mâinii. - la laba. Dezvoltarea blastemului transplantat în partea corespunzătoare a membrului anterior are loc în conformitate cu determinarea acestuia, care este determinată de nivelul amputației.Pe lângă interacțiunile intercelulare și inductive, care se dovedesc a fi mai puțin diverse decât în timpul morfogenezei embrionare, regenerarea. este influenţată semnificativ de reglarea nervoasă şi umorală. Acest lucru este destul de de înțeles prin faptul că regenerarea are loc într-un organism deja format, unde principalele mecanisme de reglare sunt acestea din urmă. Dintre influențele umorale, ar trebui să ne concentrăm pe acțiunea hormonilor. Aldosteronul, hormonii tiroidieni și hipofizari au un efect stimulator asupra refacerii pierderilor Orez. 8,86. Experimente de transplantare a blastemului membrului anterior în câmpul structurilor posterioare (explicații în text). Metaboliții eliberați de țesutul deteriorat și transportați de plasma sanguină sau transmisi prin lichidul intercelular au un efect similar. Acesta este motivul pentru care deteriorarea suplimentară în unele cazuri accelerează procesul de regenerare. Pe lângă cele de mai sus, regenerarea este influențată și de alți factori, printre care temperatura la care are loc recuperarea, vârsta animalului, funcționarea organului care stimulează regenerarea și, în anumite situații, o modificare a sarcinii electrice în regenera. S-a stabilit că schimbări reale în activitatea electrică apar la membrele amfibienilor după amputare și în timpul procesului de regenerare. Când un curent electric trece printr-un membru amputat, broaștele adulte cu gheare prezintă o regenerare crescută a membrelor anterioare. În regenerează, cantitatea de țesut nervos crește, din care se concluzionează că curentul electric stimulează creșterea în interior a nervilor în marginile membrelor, care în mod normal nu se regenerează. Încercările de a stimula regenerarea membrelor la mamifere într-un mod similar au fost fără succes. Sub influența unui curent electric sau prin combinarea acțiunii unui curent electric cu un factor de creștere nervoasă, s-a putut obține la șobolani doar creșterea țesutului scheletic sub formă de calusuri cartilaginoase și osoase, care nu semănau cu elemente normale. a scheletului membrelor. Una dintre cele mai interesante din teoria regenerării este problema surselor sale celulare. De unde provin celulele blastemale nediferențiate, similare morfologic cu celulele mezenchimale sau cum apar ele? În prezent sunt trei posibilesurse de regenerare.Primul estecelule dediferențiateal doilea -celule stem regionaleiar al treilea -celule stem din alte structuri,migrat la locul de regenerare. Majoritatea cercetătorilor recunosc dediferențierea și metaplazia în timpul regenerării cristalinului la amfibieni. Semnificația teoretică a acestei probleme constă în presupunerea posibilității sau imposibilității ca o celulă să-și schimbe programul într-o asemenea măsură încât să revină la o stare în care este din nou capabilă să-și împartă și să reprogrameze aparatul sintetic. Prezența celulelor stem regionale a fost acum stabilită în multe țesuturi: mușchi, oase, epiderma pielii, ficat, retină și altele. Astfel de celule se găsesc chiar și în țesutul nervos - în anumite zone ale creierului. În multe cazuri, se crede că acestea sunt sursa din care se formează celule diferențiate în timpul regenerării (medicina regenerativă, medicina veterinară regenerativă). Se presupune că pe măsură ce vârsta unui individ crește, populația de celule stem regionale scade. Dacă unui organ îi lipsesc propriile celule stem regionale, atunci celulele altora pot migra în el și pot da naștere țesutului dorit. S-a demonstrat recent că celulele stem izolate dintr-un țesut adult pot da naștere la celule mature din alte linii celulare, indiferent de scopul stratului germinativ clasic. Astfel, endoteliul arterelor principale mari nu are propriile rezerve de celule stem. Reînnoirea sa are loc datorită pătrunderii celulelor stem din măduva osoasă în fluxul sanguin. Cu toate acestea, ineficacitatea comparativă a unor astfel de transformări in vivo(în organism), chiar și în prezența unei leziuni tisulare, ridică întrebarea dacă acest mecanism are semnificație fiziologică. Interesant, printre celulele stem adulte, capacitatea de a schimba descendența este cea mai mare în celulele stem care pot fi cultivate în mediu pentru mult timp Dacă problema transformării liniilor celulare poate fi rezolvată, atunci va fi foarte posibil să se utilizeze aceste tehnologii în medicina reparatorie pentru a trata o gamă largă de boli. Cu toate acestea, în ciuda realizărilor biologiei din ultimii ani, există încă multe probleme nerezolvate în problema regenerării.
Regenerarea (în patologie) este refacerea integrității țesuturilor deteriorate de orice proces de boală sau influență traumatică externă. Recuperarea are loc datorită celulelor învecinate, umplerea defectului cu celule tinere și transformarea lor ulterioară în țesut matur. Această formă se numește regenerare reparatorie (compensatorie). În acest caz, sunt posibile două opțiuni de regenerare: 1) pierderea este compensată de țesut de același tip cu cel care a murit (regenerare completă); 2) pierderea este înlocuită cu țesut conjunctiv tânăr (de granulare), care se transformă în țesut cicatricial (regenerare incompletă), care nu este regenerare în sensul propriu, ci vindecarea unui defect de țesut.
Regenerarea este precedată de eliberarea unei anumite zone din celulele moarte prin topire enzimatică și absorbție în limfă sau sânge sau prin (vezi). Produsele de topire sunt unul dintre stimulatorii proliferării celulelor învecinate. În multe organe și sisteme există zone ale căror celule sunt o sursă de proliferare celulară în timpul regenerării. De exemplu, în sistemul osos o astfel de sursă este periostul, ale cărui celule, atunci când se înmulțesc, formează mai întâi țesut osteoid, care ulterior se transformă în os; în membranele mucoase - celule ale glandelor adânci (cripte). Regenerarea celulelor sanguine are loc în măduva osoasă și în afara acesteia în sistem și derivații săi (ganglioni limfatici, splină).
Nu toate țesuturile au capacitatea de a se regenera și nu în aceeași măsură. Astfel, celulele musculare ale inimii nu sunt capabile de reproducere, ducând la formarea fibrelor musculare mature, prin urmare orice defect al mușchilor miocardici este înlocuit cu o cicatrice (în special, după un atac de cord). Când țesutul cerebral moare (după hemoragie, înmuiere arteriosclerotică), defectul nu este înlocuit cu țesut nervos, ci se formează un țesut.
Uneori, țesutul care apare în timpul regenerării diferă ca structură de cel original (regenerare atipică) sau volumul său depășește volumul țesutului mort (hiperregenerare). Acest curs al procesului de regenerare poate duce la creșterea tumorii.
Regenerare (în latină regenerate - revival, restaurare) - refacerea integrității anatomice a unui organ sau țesut după moartea elementelor structurale.
În condiții fiziologice, procesele de regenerare au loc continuu cu intensitate variabilă în diferite organe și țesuturi, în funcție de intensitatea îmbătrânirii elementelor celulare ale unui anumit organ sau țesut și înlocuirea acestora cu altele nou formate. Elementele formate din sânge, celulele epiteliului tegumentar al pielii, membranele mucoase ale tractului gastrointestinal și tractul respirator sunt înlocuite în mod continuu. Procesele ciclice din sistemul reproductiv feminin duc la respingerea ritmică și reînnoirea endometrului prin regenerarea acestuia.
Toate aceste procese sunt prototipul fiziologic al regenerării patologice (se mai numește și reparatorie). Caracteristicile dezvoltării, cursului și rezultatului regenerării reparatorii sunt determinate de amploarea morții țesuturilor și de natura influențelor patogene. Trebuie avută în vedere în special ultima împrejurare, deoarece condițiile și cauzele morții tisulare sunt esențiale pentru procesul de regenerare și rezultatele acestuia. De exemplu, cicatricile după arsurile pielii au un caracter aparte, diferit de cicatricile de alte origini; cicatricile sifilitice sunt aspre, duc la retractii profunde si desfigurarea organului etc. Spre deosebire de regenerarea fiziologica, regenerarea reparatorie acoperă o gamă largă de procese care conduc la compensarea defectului cauzat de pierderea de țesut din cauza deteriorarii acestuia. Se face distincție între regenerarea reparatorie completă - restituire (înlocuirea defectului cu țesut de același tip și aceeași structură cu cel mort) și regenerarea reparatorie incompletă (umplerea defectului cu țesut care are proprietăți plastice mai mari decât cel mort). , adică țesut de granulație obișnuit și țesut conjunctiv cu transformarea în continuare în țesut cicatricial). Astfel, în patologie, regenerarea înseamnă adesea vindecare.
Conceptul de regenerare este asociat și cu conceptul de organizare, deoarece ambele procese se bazează pe legile generale ale formării de țesuturi noi și pe conceptul de substituție, adică deplasarea și înlocuirea țesutului preexistent cu țesut nou format (de exemplu, substituția). a unui cheag de sânge cu țesut fibros).
Gradul de completitudine al regenerării este determinat de doi factori principali: 1) potențialul de regenerare al unui țesut dat; 2) volumul defectului și aceeași specie sau specii eterogene de țesut mort.
Primul factor este adesea asociat cu gradul de diferențiere a unui țesut dat. Cu toate acestea, însuși conceptul de diferențiere și conținutul acestui concept sunt foarte relative, iar compararea țesuturilor pe această bază cu stabilirea unei gradații cantitative a diferențierii în termeni funcționali și morfologici este imposibilă. Alături de țesuturile care au un potențial de regenerare ridicat (de exemplu, țesut hepatic, mucoase ale tractului gastrointestinal, organe hematopoietice etc.), există organe cu un potențial nesemnificativ de regenerare, în care regenerarea nu se termină niciodată cu refacerea completă a pierdutului. țesut (de exemplu, miocard, SNC). Țesutul conjunctiv, elementele de perete ale celor mai mici vase sanguine și limfatice, nervii periferici, țesutul reticular și derivații săi au plasticitate extrem de ridicată. Prin urmare, iritația plastică, care este o traumă în sensul larg al cuvântului (adică toate formele sale), stimulează în primul rând creșterea acestor țesuturi.
Volumul de țesut mort este esențial pentru integralitatea regenerării, iar limitele cantitative ale pierderii de țesut pentru fiecare organ, care determină gradul de restaurare, sunt mai mult sau mai puțin cunoscute empiric. Se crede că pentru completitatea regenerării, nu numai volumul ca categorie pur cantitativă este important, ci și diversitatea complexă a țesuturilor moarte (aceasta se aplică în special morții tisulare cauzate de influențe toxic-infecțioase). Pentru a explica acest fapt, ar trebui, aparent, să ne întoarcem la modelele generale de stimulare a proceselor plastice în condiții patologice: stimulatorii sunt înșiși produsele morții tisulare („necrohormoni”, „razele mitogenetice”, „trefonii” etc.) ipotetici. ). Unele dintre ele sunt stimulatoare specifice pentru celule de un anumit tip, altele sunt nespecifice, stimulând cele mai multe țesuturi plastice. Stimulantii nespecifici includ produse ale defalcării și activității vitale a leucocitelor. Prezența lor în timpul inflamației reactive, care se dezvoltă întotdeauna odată cu moartea nu numai a elementelor parenchimatoase, ci și a stromei vasculare, favorizează proliferarea celor mai multe elemente plastice - țesutul conjunctiv, adică eventuala dezvoltare a unei cicatrici.
Există o schemă generală pentru succesiunea proceselor de regenerare, indiferent de zona în care are loc. În condiții patologice, procesele de regenerare în sensul restrâns al cuvântului și procesele de vindecare sunt de altă natură. Această diferență este determinată de natura morții tisulare și de direcția selectivă de acțiune a factorului patogen. Formele pure de regenerare, adică refacerea țesutului identic cu cel pierdut, se observă în cazurile în care doar anumite elemente parenchimatoase ale unui organ mor sub influența influenței patogene, cu condiția să aibă o potență regenerativă ridicată. Un exemplu în acest sens este regenerarea epiteliului tubular renal afectat selectiv prin expunerea toxică; regenerarea epiteliului mucoaselor în timpul descuamării; regenerarea alveolocitelor pulmonare în catarul descuamativ; regenerarea epiteliului pielii; regenerarea endoteliului vaselor de sânge și a endocardului etc. În aceste cazuri, sursa de regenerare o constituie elementele celulare rămase, a căror reproducere, maturare și diferențiere duce la înlocuirea completă a elementelor parenchimatoase pierdute. Când complexele structurale complexe mor, refacerea țesutului pierdut are loc din zone speciale ale organului, care sunt centre unice de regenerare. În mucoasa intestinală, în endometru, astfel de centre sunt cripte glandulare. Celulele lor înmulțite acoperă defectul mai întâi cu un strat de celule nediferențiate, din care apoi glandele se diferențiază și structura mucoasei este restaurată. În sistemul osos, un astfel de centru de regenerare este periostul, în epiteliul scuamos tegumentar - stratul Malpighian, în sistemul sanguin - măduva osoasă și derivații extramedulari ai țesutului reticular.
Legea generală a regenerării este legea dezvoltării, conform căreia, în procesul de neoplasm, iau naștere derivați celulari tineri nediferențiați, care trec ulterior prin etape de diferențiere morfologică și funcțională până la formarea țesutului matur.
Moartea zonelor unui organ constând dintr-un complex de diferite țesuturi provoacă inflamație reactivă (vezi) de-a lungul periferiei. Acesta este un act adaptativ, deoarece reacția inflamatorie este însoțită de hiperemie și creșterea metabolismului tisular, care favorizează creșterea celulelor nou formate. În plus, elementele celulare inflamatorii din grupul de histofagocite sunt material plastic pentru formarea țesutului conjunctiv.
În patologie, vindecarea anatomică este adesea realizată cu ajutorul țesutului de granulație (vezi) - stadiul formării noi a unei cicatrici fibroase. Țesutul de granulație se dezvoltă în timpul aproape oricărei regenerări reparatorii, dar gradul de dezvoltare și rezultatele finale variază în limite foarte largi. Uneori, acestea sunt zone sensibile de țesut fibros care sunt greu de distins în timpul examinării microscopice, uneori sunt fire grosiere dense de țesut cicatricial braditrofic hialinizat, adesea supuse calcificării (vezi) și osificării.
Pe lângă potențialul de regenerare al unui țesut dat, natura leziunilor sale, volumul acestuia, factorii generali sunt importanți în procesul de regenerare. Acestea includ vârsta subiectului, natura și caracteristicile nutriției și reactivitatea generală a corpului. În caz de tulburări de inervație sau deficiențe de vitamine, cursul obișnuit al regenerării reparatorii este distorsionat, ceea ce se exprimă cel mai adesea într-o încetinire a procesului de regenerare și lentă a reacțiilor celulare. Există, de asemenea, conceptul de diateză fibroplastică ca trăsătură constituțională a organismului pentru a răspunde la diferite iritații patogene cu formarea crescută de țesut fibros, care se manifestă prin formarea de cheloid (vezi), boală adezivă. În practica clinică, este important să se țină cont de factorii generali pentru a crea condiții optime pentru completarea procesului de regenerare și vindecare.
Regenerarea este unul dintre cele mai importante procese adaptative care asigură restabilirea sănătății și continuarea vieții în circumstanțe de urgență create de boală. Cu toate acestea, ca orice proces adaptativ, regenerarea la o anumită etapă și de-a lungul anumitor căi de dezvoltare își poate pierde semnificația adaptativă și poate crea ea însăși noi forme de patologie. Cicatricile deformante care deformează un organ și îi afectează brusc funcția (de exemplu, transformarea cicatricială a valvelor cardiace ca urmare a endocarditei) creează adesea o patologie cronică severă care necesită măsuri terapeutice speciale. Uneori, țesutul nou format depășește cantitativ volumul țesutului mort (super-regenerare). În plus, în fiecare regenerare există elemente de atipie, a căror severitate ascuțită este o etapă de dezvoltare a tumorii (vezi). Regenerarea organelor și țesuturilor individuale - consultați articolele relevante despre organe și țesuturi.
Informații generale
Regenerare(din lat. regenerare - renaștere) - refacerea (înlocuirea) elementelor structurale ale țesutului pentru înlocuirea morților. În sens biologic, regenerarea este proces adaptativ dezvoltat pe parcursul evoluției și inerent tuturor viețuitoarelor. În viața unui organism, fiecare funcție funcțională necesită cheltuirea unui substrat material și restaurarea acestuia. Prin urmare, în timpul regenerării există auto-reproducerea materiei vii, Mai mult, această auto-reproducere a celor vii reflectă principiul autoreglăriiȘi automatizarea funcțiilor vitale(Davydovsky I.V., 1969).
Restaurarea regenerativă a structurii poate avea loc la diferite niveluri - molecular, subcelular, celular, tisular și de organ, dar vorbim întotdeauna despre înlocuirea unei structuri care este capabilă să îndeplinească o funcție specializată. Regenerarea este restaurarea atât a structurii cât și a funcției. Semnificația procesului de regenerare constă în suportul material al homeostaziei.
Restaurarea structurii și funcției poate fi efectuată folosind procese hiperplazice celulare sau intracelulare. Pe această bază, se disting formele celulare și intracelulare de regenerare (Sarkisov D.S., 1977). Pentru formă celulară regenerarea se caracterizează prin reproducerea celulară pe cale mitotică şi amitotică, pt forma intracelulara, care pot fi organoide și intraorganoide - o creștere a numărului (hiperplaziei) și mărimii (hipertrofiei) ultrastructurilor (nuclei, nucleoli, mitocondrii, ribozomi, complex lamelar etc.) și a componentelor acestora (vezi Fig. 5, 11, 15) . Forma intracelulară regenerarea este universal, întrucât este caracteristic tuturor organelor și țesuturilor. Cu toate acestea, specializarea structurală și funcțională a organelor și țesuturilor în filo- și ontogeneză a „selectat” pentru unii forma predominant celulară, pentru alții - predominant sau exclusiv intracelular, pentru alții - ambele forme de regenerare în mod egal (Tabelul 5). Predominanța uneia sau alteia forme de regenerare în anumite organe și țesuturi este determinată de scopul lor funcțional, specializarea structurală și funcțională. Necesitatea păstrării integrității tegumentului corpului explică, de exemplu, predominanța formei celulare de regenerare a epiteliului atât a pielii, cât și a membranelor mucoase. Funcția specializată a celulei piramidale a creierului
creierul, precum și celula musculară a inimii, exclude posibilitatea divizării acestor celule și face posibilă înțelegerea necesității selecției în filo- și ontogeneză a regenerării intracelulare ca singura formă de restaurare a acestui substrat.
Tabelul 5. Forme de regenerare în organele și țesuturile mamiferelor (conform lui Sarkisov D.S., 1988)
Aceste date resping ideile care existau până de curând despre pierderea capacității unor organe și țesuturi de mamifere de a se regenera, despre regenerarea „prost” și „bine” a țesuturilor umane și ideea că există o „lege a relației inverse” între gradul de diferențiere a țesuturilor și capacitatea lor de a se regenera . S-a stabilit acum că în cursul evoluției, capacitatea de a se regenera în unele țesuturi și organe nu a dispărut, ci a luat forme (celulare sau intracelulare) corespunzătoare originalității lor structurale și funcționale (Sarkisov D.S., 1977). Astfel, toate țesuturile și organele au capacitatea de a se regenera; doar formele sale diferă în funcție de specializarea structurală și funcțională a țesutului sau organului.
Morfogeneza Procesul de regenerare constă din două faze - proliferare și diferențiere. Aceste faze sunt deosebit de bine exprimate în forma celulară de regenerare. ÎN faza de proliferare celulele tinere, nediferențiate se înmulțesc. Aceste celule sunt numite cambial(din lat. cambium- schimb, schimbare), celule stemȘi celule progenitoare.
Fiecare țesut este caracterizat de propriile celule cambiale, care diferă prin gradul de activitate proliferativă și specializare, cu toate acestea, o celulă stem poate fi strămoșul mai multor specii.
celule (de exemplu, celule stem ale sistemului hematopoietic, țesut limfoid, unii reprezentanți celulari ai țesutului conjunctiv).
ÎN faza de diferentiere celulele tinere se maturizează şi are loc specializarea lor structurală şi funcţională. Aceeași schimbare de la hiperplazia ultrastructurilor la diferențierea (maturarea) acestora stă la baza mecanismului de regenerare intracelulară.
Reglarea procesului de regenerare. Mecanismele de reglare ale regenerării includ umoral, imunologic, nervos și funcțional.
Mecanisme umorale sunt implementate atât în celulele organelor și țesuturilor lezate (regulatori intratissuale și intracelulari), cât și în afara acestora (hormoni, poetine, mediatori, factori de creștere etc.). Regulatorii umorali includ Keylons (din greaca chalaino- slabesc) - substante care pot suprima diviziunea celulara si sinteza ADN-ului; sunt specifice țesuturilor. Mecanisme imunologice reglementările sunt asociate cu „informații regenerative” transportate de limfocite. În acest sens, trebuie menționat că mecanismele homeostaziei imunologice determină și homeostazia structurală. Mecanisme nervoase procesele regenerative sunt asociate în primul rând cu funcția trofică a sistemului nervos și mecanisme funcționale- cu „cererea” funcțională a unui organ sau țesut, care este considerată ca un stimul pentru regenerare.
Dezvoltarea procesului de regenerare depinde în mare măsură de o serie de condiții sau factori generali și locali. LA general ar trebui să includă vârsta, constituția, starea nutrițională, starea metabolică și hematopoietică, local - starea de inervație, circulația sanguină și limfatică a țesutului, activitatea proliferativă a celulelor acestuia, natura procesului patologic.
Clasificare. Există trei tipuri de regenerare: fiziologică, reparatorie și patologică.
Regenerare fiziologică apare pe tot parcursul vieții și se caracterizează prin reînnoirea constantă a celulelor, a structurilor fibroase și a substanței de bază a țesutului conjunctiv. Nu există structuri care să nu sufere regenerare fiziologică. Acolo unde domină forma celulară de regenerare, are loc reînnoirea celulară. Astfel, există o schimbare constantă a epiteliului tegumentar al pielii și mucoaselor, a epiteliului secretor al glandelor exocrine, a celulelor care căptușesc membranele seroase și sinoviale, a elementelor celulare ale țesutului conjunctiv, a celulelor roșii din sânge, a leucocitelor și a trombocitelor, etc. În țesuturile și organele în care se pierde forma celulară de regenerare, de exemplu în inimă, creier, structurile intracelulare sunt reînnoite. Odată cu reînnoirea celulelor și a structurilor subcelulare, regenerare biochimica, acestea. reînnoirea compoziției moleculare a tuturor componentelor corpului.
Regenerare reparatoare sau restauratoare observat în diferite procese patologice care duc la deteriorarea celulelor și țesutului
a ei. Mecanismele de regenerare reparatoare și fiziologică sunt aceleași; regenerarea reparatorie este regenerarea fiziologică îmbunătățită. Cu toate acestea, datorită faptului că regenerarea reparatorie este stimulată de procese patologice, are diferențe morfologice calitative față de cele fiziologice. Regenerarea reparatorie poate fi completă sau incompletă.
Regenerare completa, sau restituire, caracterizat prin compensarea defectului cu tesut identic cu cel mort. Se dezvoltă predominant în țesuturi unde predomină regenerarea celulară. Astfel, in tesutul conjunctiv, oase, piele si membranele mucoase, chiar si defectele de organ relativ mari pot fi inlocuite prin diviziunea celulara prin tesut identic cu cel mort. La regenerare incompletă, sau substituţie, defectul este înlocuit cu țesut conjunctiv, cicatrice. Substituția este caracteristică organelor și țesuturilor în care predomină forma intracelulară de regenerare, sau este combinată cu regenerarea celulară. Deoarece regenerarea presupune refacerea unei structuri capabile să îndeplinească o funcție specializată, semnificația regenerării incomplete nu constă în înlocuirea defectului cu o cicatrice, ci în hiperplazie compensatorie elemente ale țesutului specializat rămas, a căror masă crește, adică se întâmplă hipertrofie tesaturi.
La regenerare incompletă, acestea. vindecarea țesutului cu o cicatrice, hipertrofia apare ca expresie a procesului de regenerare, motiv pentru care se numește regenerativ, conţine sensul biologic de regenerare reparatorie. Hipertrofia regenerativă poate fi realizată în două moduri - prin hiperplazie sau hiperplazie celulară și hipertrofie a ultrastructurilor celulare, adică. hipertrofie celulară.
Restaurarea masei inițiale a organului și a funcției sale datorită în primul rând hiperplazie celulară apare în timpul hipertrofiei regenerative a ficatului, rinichilor, pancreasului, glandelor suprarenale, plămânilor, splinei etc. Hipertrofia regenerativă datorată hiperplazia ultrastructurilor celulare caracteristic miocardului, creierului, i.e. acele organe în care predomină forma intracelulară de regenerare. În miocard, de exemplu, de-a lungul periferiei cicatricei care a înlocuit infarctul, dimensiunea fibrelor musculare crește semnificativ, adică. ele hipertrofiază datorită hiperplaziei elementelor lor subcelulare (Fig. 81). Ambele căi de hipertrofie regenerativă nu se exclud reciproc, ci, dimpotrivă, adesea combina. Astfel, cu hipertrofia regenerativă a ficatului, apare nu numai o creștere a numărului de celule în partea de organ conservată după lezare, ci și hipertrofia acestora, cauzată de hiperplazia ultrastructurilor. Nu se poate exclude ca în mușchiul cardiac hipertrofia regenerativă să se producă nu numai sub formă de hipertrofie a fibrelor, ci și prin creșterea numărului de celule musculare care le alcătuiesc.
Perioada de recuperare nu se limitează de obicei doar la faptul că regenerarea reparatorie se desfășoară în organul afectat. Dacă
 Orez. 81. Hipertrofia miocardică regenerativă. Fibrele musculare hipertrofiate sunt situate de-a lungul periferiei cicatricii
Orez. 81. Hipertrofia miocardică regenerativă. Fibrele musculare hipertrofiate sunt situate de-a lungul periferiei cicatricii
influența factorului patogen încetează până la moartea celulelor și are loc refacerea treptată a organelelor deteriorate. În consecință, manifestările reacției reparatorii ar trebui extinse pentru a include procesele intracelulare de restaurare în organele alterate distrofic. Opinia general acceptată despre regenerare doar ca etapă finală a procesului patologic este nejustificată. Regenerarea reparatorie nu este local, A reacție generală a corpului, acoperind diverse organe, dar realizându-se pe deplin doar într-unul sau altul dintre ele.
DESPRE regenerare patologică spun ei în cazurile în care, din anumite motive, există denaturarea procesului de regenerare, perturbarea schimbărilor de fază proliferare
și diferențiere. Regenerarea patologică se manifestă prin formarea excesivă sau insuficientă de țesut regenerant (hiper- sau hiporegenerare), precum și în transformarea în timpul regenerării unui tip de țesut în altul [metaplazie - vezi. Procese de ajustare (adaptare) și compensare]. Exemplele includ hiperproducția de țesut conjunctiv cu formarea cheloid, regenerarea excesivă a nervilor periferici și formarea excesivă a calusului în timpul vindecării fracturilor, vindecarea lentă a rănilor și metaplazia epitelială în focarul inflamației cronice. Regenerarea patologică se dezvoltă de obicei când încălcări ale generaleȘi condiţiile locale de regenerare(inervație afectată, foamete de proteine și vitamine, inflamație cronică etc.).
Regenerarea țesuturilor și organelor individuale
Regenerarea reparatorie a sângelui diferă de regenerarea fiziologică în primul rând prin intensitatea sa mai mare. În acest caz, măduva osoasă roșie activă apare în oasele lungi în locul măduvei osoase grase (transformarea mieloidă a măduvei osoase grase). Celulele adipoase sunt înlocuite cu insule în creștere de țesut hematopoietic, care umple canalul medular și arată suculent și roșu închis. În plus, hematopoieza începe să apară în afara măduvei osoase - extramedulară, sau extramedulară, hematopoieza. Ocha-
gi ale hematopoiezei extramedulare (heterotopice) ca urmare a evacuării celulelor stem din măduva osoasă apar în multe organe și țesuturi - splină, ficat, ganglioni limfatici, mucoase, țesut adipos etc.
Regenerarea sângelui poate fi puternic deprimat (de exemplu, cu radiații, anemie aplastică, aleukie, agranulocitoză) sau pervertit (de exemplu, cu anemie pernicioasă, policitemie, leucemie). În acest caz, elementele formate imature, inferioare funcțional și deteriorate rapid intră în sânge. În astfel de cazuri vorbim despre regenerare patologică a sângelui.
Capacitățile reparatorii ale organelor sistemului hematopoietic și imunocompetente sunt ambigue. Măduvă osoasă are proprietăți plastice foarte înalte și poate fi restaurat chiar și cu daune semnificative. Ganglionii limfatici se regenerează bine numai în cazurile în care se păstrează conexiunile vaselor limfatice aferente și eferente cu țesutul conjunctiv din jur. Regenerarea țesuturilor splină atunci când este deteriorat, este de obicei incomplet; țesutul mort este înlocuit cu o cicatrice.
Regenerarea vaselor de sânge și limfatice procedează ambiguu în funcţie de calibrul lor.
Microvase au o capacitate de regenerare mai mare decât vasele mari. O nouă formare a microvaselor poate avea loc prin înmugurire sau autogen. În timpul regenerării vasculare prin înmugurire (Fig. 82) proeminențe laterale apar în peretele lor din cauza celulelor endoteliale care se divizează rapid (angioblaste). Se formează fire de endoteliu, în care apar goluri și sângele sau limfa curge în ele din vasul „matern”. Alte elemente: peretele vascular se formează datorită diferențierii endoteliului și a celulelor țesutului conjunctiv din jurul vasului.Fibrele nervoase din nervii preexistenți cresc în peretele vascular. Neoplasm autogen vaselor este că în țesutul conjunctiv apar focare de celule nediferențiate. În aceste focare apar crăpături în care se deschid capilarele preexistente și curge sângele. Celulele tinere ale țesutului conjunctiv, diferențiate, formează căptușeala endotelială și alte elemente ale peretelui vasului.
 Orez. 82. Regenerare vasculară prin înmugurire
Orez. 82. Regenerare vasculară prin înmugurire
Vase mari nu au suficiente proprietăți plastice. Prin urmare, dacă pereții lor sunt deteriorați, doar structurile învelișului interior, căptușeala sa endotelială, sunt restaurate; elementele membranelor medii și exterioare sunt de obicei înlocuite cu țesut conjunctiv, ceea ce duce adesea la îngustarea sau obliterarea lumenului vasului.
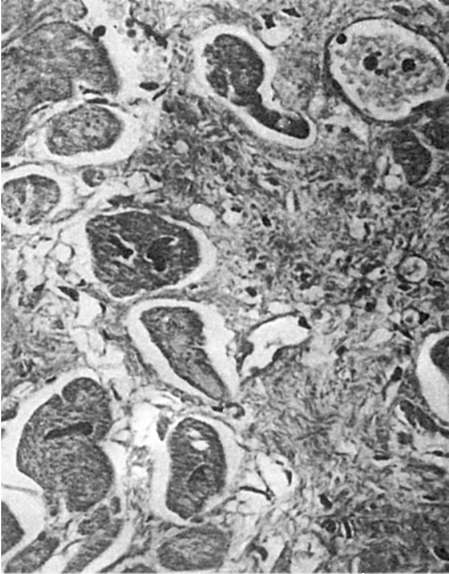
Regenerarea țesutului conjunctivîncepe cu proliferarea elementelor mezenchimatoase tinere și formarea de noi microvase. Se formează țesut conjunctiv tânăr, bogat în celule și vase cu pereți subțiri, care are un aspect caracteristic. Aceasta este o țesătură suculentă roșu închis, cu o suprafață granulară, ca și cum ar fi presărată cu granule mari, care a stat la baza denumirii acesteia. țesut de granulație. Granulele sunt bucle de vase cu pereți subțiri nou formate care ies deasupra suprafeței, care formează baza țesutului de granulație. Între vase există multe celule nediferențiate de țesut conjunctiv asemănător limfocitelor, leucocite, plasmocite și mastocite (Fig. 83). Ce se întâmplă în continuare este maturare țesut de granulație, care se bazează pe diferențierea elementelor celulare, a structurilor fibroase și a vaselor de sânge. Numărul de elemente hematogene scade, iar fibroblastele cresc. În legătură cu sinteza colagenului de către fibroblasti, argirofil(vezi Fig. 83) și apoi fibre de colagen. Sinteza glicozaminoglicanilor de către fibroblaste servește la formare
substanta principala țesut conjunctiv. Pe măsură ce fibroblastele se maturizează, numărul de fibre de colagen crește și ele sunt grupate în mănunchiuri; În același timp, numărul de vase scade, ele se diferențiază în artere și vene. Maturarea țesutului de granulație se încheie cu formarea țesut cicatricial fibros grosier.
Noua formare de țesut conjunctiv are loc nu numai atunci când este deteriorat, ci și atunci când alte țesuturi sunt regenerate incomplet, precum și în timpul organizării (încapsulării), vindecării rănilor și inflamației productive.
Maturarea țesutului de granulație poate avea anumite abaterile. Inflamația care se dezvoltă în țesutul de granulație duce la o întârziere a maturării acestuia,
 Orez. 83.Țesut de granulație. Între vasele cu pereți subțiri există multe celule de țesut conjunctiv nediferențiate și fibre argirofile. impregnare cu argint
Orez. 83.Țesut de granulație. Între vasele cu pereți subțiri există multe celule de țesut conjunctiv nediferențiate și fibre argirofile. impregnare cu argint
iar activitatea sintetică excesivă a fibroblastelor duce la formarea excesivă a fibrelor de colagen, urmată de hialinoză pronunțată. În astfel de cazuri, țesutul cicatricial apare sub forma unei forme tumorale de culoare roșie-albăstruie, care se ridică deasupra suprafeței pielii sub forma cheloid. Cicatricile cheloide se formează după diferite leziuni traumatice ale pielii, în special după arsuri.
Regenerarea țesutului adipos apare din cauza formării noi a celulelor țesutului conjunctiv, care se transformă în celule adipoase (adipocite) prin acumularea de lipide în citoplasmă. Celulele adipoase sunt pliate în lobuli, între care există straturi de țesut conjunctiv cu vase și nervi. Regenerarea țesutului adipos poate apărea și din resturile nucleate ale citoplasmei celulelor adipoase.
Regenerarea țesutului ososîn cazul fracturii osoase, depinde în mare măsură de gradul de distrugere osoasă, repoziţionarea corectă a fragmentelor osoase, condiţiile locale (afecţiuni circulatorii, inflamaţii etc.). La necomplicat poate apărea fractură osoasă, când fragmentele osoase sunt imobile unire osoasă primară(Fig. 84). Începe cu creșterea în interior a elementelor și vaselor mezenchimale tinere în zona defectului și a hematomului între fragmentele osoase. Există un așa-zis calus preliminar al țesutului conjunctiv,în care începe imediat formarea osului. Este asociat cu activarea și proliferarea osteoblasteleîn zona afectată, dar în primul rând în periostat și endostat. În țesutul fibroreticular osteogen apar grinzi osoase ușor calcificate, numărul cărora crește.
Format calus preliminar. Ulterior, se maturizează și se transformă în os lamelar matur - așa
 Orez. 84. Fuziunea osoasă primară. Calus intermediar (indicat cu o săgeată), fuziunea fragmentelor osoase (conform lui G.I. Lavrishcheva)
Orez. 84. Fuziunea osoasă primară. Calus intermediar (indicat cu o săgeată), fuziunea fragmentelor osoase (conform lui G.I. Lavrishcheva)
calus final, care prin structura sa se deosebeşte de ţesutul osos doar prin aranjarea aleatorie a traverselor osoase. După ce osul începe să-și îndeplinească funcția și apare o sarcină statică, țesutul nou format suferă o restructurare cu ajutorul osteoclastelor și osteoblastelor, apare măduva osoasă, se restabilește vascularizația și inervația. În cazul în care condițiile locale pentru regenerarea osoasă sunt încălcate (tulburări circulatorii), apar mobilitatea fragmentelor, fracturile diafizare extinse. fuziunea osoasă secundară(Fig. 85). Acest tip de fuziune osoasă se caracterizează prin formarea între fragmentele osoase ale primului țesut cartilaginos, pe baza căruia se construiește țesutul osos. Prin urmare, cu fuziunea osoasă secundară se vorbește calus osteocondral preliminar, care în cele din urmă se dezvoltă în os matur. Fuziunea osoasă secundară, în comparație cu fuziunea primară, este mult mai frecventă și durează mai mult.
La conditii nefavorabile regenerarea osoasă poate fi afectată. Astfel, atunci când o rană se infectează, regenerarea osoasă este întârziată. Fragmentele osoase, care în cursul normal al procesului de regenerare servesc drept cadru pentru țesutul osos nou format, în condiții de supurație a plăgii susțin inflamația, care inhibă regenerarea. Uneori, calusul osteocondral primar nu se diferențiază într-un calus osos. În aceste cazuri, capetele osului rupt rămân mobile și a falsă articulație. Producția excesivă de țesut osos în timpul regenerării duce la apariția pintenilor ososi - exostoze.
Regenerarea țesutului cartilajului spre deosebire de os, de obicei apare incomplet. Doar defecte mici pot fi înlocuite cu țesut nou format datorită elementelor cambiale ale pericondrului - condroblaste. Aceste celule creează substanța de bază a cartilajului și apoi se dezvoltă în celule de cartilaj matur. Defectele mari ale cartilajului sunt înlocuite cu țesut cicatricial.
Regenerarea țesutului muscular, capabilitățile și formele sale variază în funcție de tipul de țesătură. Neted Mușchii, ale căror celule au capacitatea de a suferi mitoză și amitoză, se pot regenera destul de complet cu defecte minore. Zonele semnificative de afectare a mușchilor netezi sunt înlocuite cu cicatrice, în timp ce fibrele musculare rămase suferă hipertrofie. O nouă formare a fibrelor musculare netede poate apărea prin transformarea (metaplazia) elementelor de țesut conjunctiv. Așa se formează mănunchiuri de fibre musculare netede în aderențe pleurale, în trombi în curs de organizare și în vase în timpul diferențierii lor.
Striat mușchii se regenerează numai dacă se păstrează sarcolema. În interiorul tuburilor din sarcolemă are loc regenerarea organelelor sale, rezultând apariția unor celule numite mioblaste. Se alungesc, numărul de nuclee din ele crește, în sarcoplasmă
 Orez. 85. Fuziunea osoasă secundară (conform lui G.I. Lavrishcheva):
Orez. 85. Fuziunea osoasă secundară (conform lui G.I. Lavrishcheva):
a - calus periostal osteocondral; o secțiune de țesut osos între țesutul cartilaginos (imagine microscopică); b - calus osteocondral periostal (histotopograma la 2 luni de la operatie): 1 - partea osoasa; 2 - partea cartilaginoasă; 3 - fragmente osoase; c - calus periostal, fuzionand fragmente osoase deplasate
miofibrilele se diferențiază, iar tuburile sarcolemale se transformă în fibre musculare striate. Regenerarea mușchilor scheletici poate fi, de asemenea, asociată cu celule satelit, care sunt situate sub sarcolemă, adică. în interiorul fibrei musculare și sunt cambial.În caz de rănire, celulele satelit încep să se dividă rapid, apoi suferă diferențiere și asigură refacerea fibrelor musculare. Dacă, atunci când un mușchi este deteriorat, integritatea fibrelor este perturbată, atunci apar proeminențe în formă de balon la capetele rupurilor lor, care conțin un număr mare de nuclei și se numesc rinichi musculari.În acest caz, restabilirea continuității fibrei nu are loc. Locul de ruptură este umplut cu țesut de granulație, care se transformă într-o cicatrice (calus muscular). Regenerare muschii inimii dacă este deteriorat, ca și în cazul afectarii mușchilor striați, se termină cu cicatrizarea defectului. Cu toate acestea, în fibrele musculare rămase, apare hiperplazia intensă a ultrastructurilor, ceea ce duce la hipertrofia fibrelor și restabilirea funcției organelor (vezi Fig. 81).
Regenerare epitelială se desfășoară în majoritatea cazurilor destul de complet, deoarece are o capacitate mare de regenerare. Regenerează deosebit de bine epiteliul de acoperire. Recuperare epiteliu stratificat stratificat cheratinizant scuamos posibil chiar și cu defecte ale pielii destul de mari. În timpul regenerării epidermei la marginile defectului are loc o proliferare crescută a celulelor stratului germinal (cambial) și germinal (malpighian). Celulele epiteliale rezultate acoperă mai întâi defectul într-un singur strat. Ulterior, stratul de epiteliu devine multistratificat, celulele sale se diferențiază și capătă toate semnele epidermei, inclusiv stratul germinal, granular, strălucitor (pe tălpile și suprafața palmară a mâinilor) și stratul cornos. Când regenerarea epiteliului pielii este afectată, se formează ulcere nevindecatoare, adesea cu creșterea epiteliului atipic la marginile lor, care pot servi ca bază pentru dezvoltarea cancerului de piele.
Epiteliul de acoperire al membranelor mucoase (squamous multistratificat nekeratinizat, tranzițional, monostrat prismatic și multinucleat ciliated) se regenerează în același mod ca multistratificat scuamos keratinizat. Defectul mucoasei este restabilit datorită proliferării celulelor care căptușesc criptele și canalele excretoare ale glandelor. Celulele epiteliale aplatizate nediferențiate acoperă mai întâi defectul cu un strat subțire (Fig. 86), apoi celulele capătă forma caracteristică structurilor celulare ale căptușelii epiteliale corespunzătoare. În paralel, glandele membranei mucoase (de exemplu, glandele tubulare ale intestinului, glandele endometriale) sunt parțial sau complet restaurate.
Regenerarea mezoteliului peritoneul, pleura și sacul pericardic se realizează prin divizarea celulelor supraviețuitoare. Pe suprafața defectului apar celule cubice relativ mari, care apoi se aplatizează. Pentru defecte mici, căptușeala mezotelială este restaurată rapid și complet.
Starea țesutului conjunctiv subiacent este importantă pentru refacerea epiteliului tegumentar și a mezoteliului, deoarece epitelizarea oricărui defect este posibilă numai după umplerea acestuia cu țesut de granulație.
Regenerarea epiteliului de organe specializate(ficat, pancreas, rinichi, glande endocrine, alveole pulmonare) se efectuează în funcție de tipul hipertrofie regenerativă:în zonele afectate, țesutul este înlocuit cu o cicatrice, iar de-a lungul periferiei sale apar hiperplazia și hipertrofia celulelor parenchimului. ÎN ficat zona de necroză este întotdeauna supusă cicatricilor, dar în restul organului există o formare intensă de celule noi, precum și o hiperplazie a structurilor intracelulare, care este însoțită de hipertrofia lor. Ca urmare, masa și funcția originală a organului sunt rapid restaurate. Capacitățile de regenerare ale ficatului sunt aproape nelimitate. În pancreas, procesele regenerative sunt bine exprimate atât în secțiunile exocrine, cât și în insulele pancreatice, iar epiteliul glandelor exocrine devine sursa refacerii insulelor. ÎN rinichi cu necroza epiteliului tubular, nefrocitele supraviețuitoare se înmulțesc și tubii sunt restaurați, dar numai când membrana bazală tubulară este conservată. Când este distrus (tubulorrexis), epiteliul nu este restaurat și tubul este înlocuit cu țesut conjunctiv. Epiteliul tubular mort nu este restaurat nici în cazul în care glomerulul vascular moare concomitent cu tubul. În acest caz, țesutul conjunctiv cicatricial crește în locul nefronului mort, iar nefronii din jur suferă hipertrofie regenerativă. În glande secretie interna procesele de restaurare sunt reprezentate şi de regenerare incompletă. ÎN plămâni după îndepărtarea lobilor individuali, în partea rămasă apare hipertrofia și hiperplazia elementelor tisulare. Regenerarea epiteliului specializat al organelor poate avea loc atipic, ceea ce duce la proliferarea țesutului conjunctiv, restructurarea structurală și deformarea organelor; în astfel de cazuri vorbim despre ciroză (ciroză hepatică, nefrociroză, pneumociroză).
Regenerarea diferitelor părți ale sistemului nervos se întâmplă în mod ambiguu. ÎN cap Și măduva spinării neoplasmele celulelor ganglionare nu pro-
 Orez. 86. Regenerarea epiteliului în partea inferioară a unui ulcer gastric cronic
Orez. 86. Regenerarea epiteliului în partea inferioară a unui ulcer gastric cronic
apare și atunci când sunt distruse, restabilirea funcției este posibilă numai prin regenerarea intracelulară a celulelor supraviețuitoare. Neuroglia, în special microglia, se caracterizează printr-o formă celulară de regenerare, prin urmare defectele țesutului creierului și măduvei spinării sunt de obicei pline de celule neurogliale în proliferare - așa-numitele glială (gliotic) cicatrici. Dacă este deteriorat nodurile vegetative Odată cu hiperplazia ultrastructurilor celulare, apare și noua lor formare. În caz de încălcare a integrităţii nervul periferic regenerarea are loc datorită segmentului central, care și-a păstrat legătura cu celula, în timp ce segmentul periferic moare. Celulele înmulțitoare ale tecii Schwann ale segmentului periferic mort al nervului sunt situate de-a lungul acestuia și formează o teacă - așa-numitul cordon Büngner, în care cresc cilindrii axiali regenerați din segmentul proximal. Regenerarea fibrelor nervoase se termină cu mielinizarea lor și refacerea terminațiilor nervoase. Hiperplazie regenerativă receptori, dispozitivele sinaptice pericelulare și efectorii sunt uneori însoțite de hipertrofia aparatului lor terminal. Dacă regenerarea nervilor este întreruptă dintr-un motiv sau altul (divergență semnificativă a unor părți ale nervului, dezvoltarea unui proces inflamator), atunci la locul întreruperii acestuia se formează o cicatrice în care cilindrii axiali regenerați ai segmentului proximal al nervului. sunt localizate aleatoriu. Creșteri similare apar la capetele nervilor tăiați în ciotul unui membru după amputare. Se numesc astfel de creșteri formate din fibre nervoase și țesut fibros neuroame de amputație.
Vindecarea ranilor
Vindecarea rănilor se desfășoară în conformitate cu legile regenerării reparatorii. Rata de vindecare a rănilor și rezultatele acesteia depind de gradul și profunzimea leziunii rănii, de caracteristicile structurale ale organului, de starea generală a corpului și de metodele de tratament utilizate. Potrivit lui I.V. Davydovsky, se disting următoarele tipuri de vindecare a rănilor: 1) închiderea directă a defectului epitelial; 2) vindecarea sub crusta; 3) vindecarea rănilor prin intenție primară; 4) vindecarea rănilor prin intenție secundară sau vindecarea rănilor prin supurație.
Închiderea directă a defectului epitelial- aceasta este cea mai simplă vindecare, care constă în târâirea epiteliului peste defectul de suprafață și acoperirea acestuia cu un strat epitelial. Se observă pe cornee, mucoase vindecarea sub crusta se referă la defecte mici, pe suprafața cărora apare rapid o crustă de uscare (crustă) de sânge coagulat și limfă; epiderma este restabilită sub o crustă, care dispare la 3-5 zile după leziune.
Vindecarea prin intenție primară (per rimam intentionem) observat în răni cu afectare nu numai a pielii, ci și a țesutului subiacent,
iar marginile rănii sunt uniforme. Rana este umplută cu cheaguri de sânge vărsat, care protejează marginile rănii de deshidratare și infecție. Sub influența enzimelor proteolitice ale neutrofilelor, are loc liza parțială a coagulării sângelui și a detritusului tisular. Neutrofilele mor și sunt înlocuite cu macrofage, care fagocitează globulele roșii și resturile de țesut deteriorat; Hemosiderina se găsește la marginile plăgii. O parte din conținutul plăgii este îndepărtată în prima zi a plăgii împreună cu exudatul independent sau în timpul tratamentului plăgii - curatare primara. În a 2-3-a zi, fibroblastele și capilarele nou formate apar la marginile plăgii, crescând unul spre celălalt, țesut de granulație, al cărui strat nu atinge dimensiuni mari în timpul tensiunii primare. Până în ziua 10-15 se maturizează complet, defectul plăgii este epitelializat și rana se vindecă cu o cicatrice delicată. Într-o plagă chirurgicală, vindecarea prin intenție primară este accelerată datorită faptului că marginile acesteia sunt strânse cu fire de mătase sau catgut, în jurul cărora se acumulează celule gigantice de corpuri străine care le absorb și nu interferează cu vindecarea.
Vindecarea prin intenție secundară (per secundam intentionem), sau vindecarea prin supurație (sau vindecarea prin granulație - pe granulare), Se observă de obicei cu răni extinse, însoțite de zdrobirea și necroza țesuturilor, pătrunderea corpurilor străine și a microbilor în rană. Hemoragiile și umflarea traumatică a marginilor plăgii apar la locul plăgii și apar rapid semne de demarcație. inflamație purulentă la limita cu tesut mort, topirea maselor necrotice. În primele 5-6 zile, masele necrotice sunt respinse - secundar curățarea plăgii, iar țesutul de granulație începe să se dezvolte la marginile plăgii. țesut de granulație, umplerea plăgii, constă din 6 straturi care trec unul în celălalt (Anichkov N.N., 1951): strat superficial leucocitar-necrotic; strat superficial de anse vasculare, strat de vase verticale, strat de maturizare, strat de fibroblaste situate orizontal, strat fibros. Maturarea țesutului de granulație în timpul vindecării rănilor cu intenție secundară este însoțită de regenerarea epitelială. Cu toate acestea, cu acest tip de vindecare a rănilor, la locul său se formează întotdeauna o cicatrice.
Regenerare- refacerea de către organism a organelor și țesuturilor pierdute sau deteriorate, precum și refacerea întregului organism din partea sa. Într-o măsură mai mare
într-o măsură mai mică – inerente plantelor și animalelor nevertebrate. Regenerarea poate fi cauzată
experimental.
Regenerare are ca scop refacerea elementelor structurale deteriorate iar procesele de regenerare pot
realizate la diferite niveluri:
a) moleculară
b) subcelular
c) celular - reproducere celulară prin mitoză şi cale amitotică
d) tesut
d) organ.
Tipuri de regenerare:
7. fiziologic - asigură funcţionarea organelor şi sistemelor în condiţii normale. Regenerarea fiziologică are loc în toate organele, dar în unele mai mult, în altele mai puțin.
2. Reparator(restaurativ) - apare în legătură cu un proces patologic care duce la deteriorarea țesuturilor (aceasta este o regenerare fiziologică îmbunătățită)
a) regenerare completă (restituire) - exact același țesut apare la locul leziunii tisulare
b) regenerare incompletă (substituţie) - în locul ţesutului mort apare ţesutul conjunctiv. De exemplu, în inimă în timpul infarctului miocardic, apare necroza, care este înlocuită cu țesut conjunctiv.
Semnificația regenerării incomplete: hipertrofia regenerativă apare în jurul țesutului conjunctiv, care
asigură păstrarea funcţiei organului afectat.
Hipertrofie regenerativă realizat prin:
a) hiperplazie celulară (formare excesivă)
b) hipertrofia celulară (creșterea volumului și greutății organelor).
Hipertrofia regenerativă a miocardului se realizează din cauza hiperplaziei structurilor intracelulare.
Forme de regenerare.
1. Celulară - reproducerea celulară are loc pe cale mitotică și amitotică. Există în țesutul osos, epidermă, mucoasa gastrointestinală, mucoasa respiratorie, mucoasa genito-urinar, endoteliu, mezoteliu, țesut conjunctiv lax, sistemul hematopoietic. Regenerarea completă are loc în aceste organe și țesuturi (exact același țesut).
2. Intracelular – apare hiperplazia structurilor intracelulare. Miocard, mușchi scheletici (în principal), celule ganglionare ale sistemului nervos central (exclusiv).
3. Forme celulare și intracelulare. Ficat, rinichi, plămâni, mușchi netezi, sistem nervos autonom, pancreas, sistem endocrin. De obicei apare regenerarea incompletă.
Regenerarea țesutului conjunctiv.
Etape:
1. Formarea țesutului de granulație. Treptat, vasele și celulele sunt deplasate odată cu formarea de fibre. Fibroblastele sunt fibrocite care produc fibre.
2. Formarea de țesut conjunctiv matur. Regenerarea sângelui
1. Regenerare fiziologică. În măduva osoasă.
2. Regenerare reparatoare. Apare cu anemie, leucopenie, trombocitopenie. Apar focare extramedulare de hematopoieză (în ficat, splină, ganglioni limfatici, măduva osoasă galbenă este implicată în hematopoieză).
3. Regenerare patologică. Pentru boala de radiații, leucemie. În organele hematopoietice, imature
elemente hematopoietice (celule de putere).
Întrebarea 16
HOMEOSTAZA.
Homeostazia – menținerea constantă a mediului intern al organismului în condiții de mediu în continuă schimbare. Deoarece organismul este un obiect autoreglabil pe mai multe niveluri, poate fi considerat din punctul de vedere al ciberneticii. Apoi, corpul este un sistem complex de autoreglare pe mai multe niveluri, cu multe variabile.
Variabile de intrare:
Cauză;
Iritarea.
Variabile de ieșire:
Reacţie;
Consecinţă.
Motivul este o abatere de la reacția normală a organismului. Feedback-ul joacă un rol decisiv. Există feedback pozitiv și negativ.
Feedback negativ reduce efectul semnalului de intrare asupra semnalului de ieșire. Feedback pozitiv crește efectul semnalului de intrare asupra efectului de ieșire al acțiunii.
Un organism viu este un sistem ultrastabil care caută cea mai optimă stare stabilă, care este asigurată de adaptări.
Întrebarea 18:
PROBLEME DE TRANSPLANT.
Transplantul este transplantul de țesuturi și organe.
Transplantul la animale și la om este grefarea de organe sau secțiuni de țesut individual pentru a înlocui defectele, a stimula regenerarea, în timpul operațiilor cosmetice, precum și în scopuri de experiment și terapie tisulară.
Autotransplantul este un transplant de țesut într-un singur organism, alotransplantul este un transplant între organisme din aceeași specie. Xenotransplantul este un transplant între diferite specii.
Întrebarea 19
Cronobiologie- o ramură a biologiei care studiază ritmurile biologice, cursul diferitelor procese biologice
(în mare parte ciclic) în timp.
Ritmuri biologice- (bioritmuri), fluctuații ciclice ale intensității și naturii proceselor și fenomenelor biologice. Unele ritmuri biologice sunt relativ independente (de exemplu, frecvența contracțiilor inimii, respirația), altele sunt asociate cu adaptarea organismelor la ciclurile geofizice - zilnic (de exemplu, fluctuații în intensitatea diviziunii celulare, metabolismul, activitatea motorie a animalelor). ), mareale (de exemplu, procese biologice în organismele asociate cu nivelul mareelor mării), anuale (modificări ale numărului și activității animalelor, creșterea și dezvoltarea plantelor etc.). Știința ritmurilor biologice este cronobiologia.
Întrebarea 20
FILGENEZA SCHELETICĂ
Scheletul de pește este format dintr-un craniu, coloana vertebrală, schelet de înotătoare nepereche și pereche și curele lor. În regiunea trunchiului, coastele sunt atașate proceselor transversale ale corpului. Vertebrele se articulează între ele folosind procese articulare, asigurând îndoirea în primul rând în plan orizontal.
Scheletul amfibienilor, ca toate vertebratele, este format din craniul, coloana vertebrală, scheletul membrelor și brâurile acestora. Craniul este aproape în întregime cartilaginos (Fig. 11.20). Este articulat mobil cu coloana vertebrală. Coloana vertebrală conține nouă vertebre, unite în trei secțiuni: cervicală (1 vertebră), trunchi (7 vertebre), sacră (1 vertebră) și toate vertebrele caudale sunt fuzionate pentru a forma un singur os - urostilul. Nu există coaste. Brâul scapular include oase tipice vertebratelor terestre: scapule pereche, oase de corb (coracoide), clavicule și stern nepereche. Are aspectul unui jumătate de inel situat în grosimea mușchilor trunchiului, adică nu este conectat la coloana vertebrală. Centura pelviană este formată din două oase pelvine, formate din trei perechi de oase iliace, ischiatice și pubiene fuzionate. Oasele iliace lungi sunt atașate proceselor transversale ale vertebrei sacrale. Scheletul membrelor libere este construit după tipul de sistem de pârghii multi-membri, conectate mobil prin articulații sferice. Ca parte a membrului anterior. distingeți umărul, antebrațul și mâna.
Corpul șopârlei este împărțit în cap, trunchi și coadă. În secțiunea trunchi gâtul este bine definit. Întregul corp este acoperit cu solzi cornosi, iar capul și burta sunt acoperite cu scute mari. Membrele șopârlei sunt bine dezvoltate și înarmate cu cinci degete cu gheare. Oasele humerusului și femurului sunt paralele cu suprafața solului, determinând corpul să se încline și să atingă solul (de unde și numele clasei). Coloana cervicală este formată din opt vertebre, prima fiind conectată mobil atât cu craniul, cât și cu cea de-a doua vertebră, ceea ce oferă capului o mai mare libertate de mișcare. Vertebrele regiunii toracolombare poartă coaste, dintre care unele sunt conectate la stern, rezultând formarea cutiei toracice. Vertebrele sacrale asigură o conexiune mai puternică cu oasele pelvine decât la amfibieni.
Structura scheletului mamiferelor este practic similară cu cea a vertebratelor terestre, dar există unele diferențe: numărul vertebrelor cervicale este constant și egal cu șapte, craniul este mai voluminos, ceea ce este asociat cu dimensiunea mai mare a creierului. Oasele craniului fuzionează destul de târziu, permițând creierului să crească pe măsură ce animalul crește. Membrele mamiferelor sunt construite după tipul cu cinci degete, caracteristic vertebratelor terestre.
Întrebarea 21
FILGENEZA SISTEMULUI CIRCULATOR
Sistemul circulator al peștelui este obturat. Inima este cu două camere, constând dintr-un atriu și un ventricul. Sângele venos din ventriculul inimii pătrunde în aorta abdominală, care îl duce la branhii, unde este îmbogățit cu oxigen și eliberat de dioxid de carbon. Sângele arterial care curge din branhii este colectat în aorta dorsală, care este situată de-a lungul corpului sub coloana vertebrală. Numeroase artere se ramifică din aorta dorsală către diferite organe ale peștilor. În ele, arterele se despart într-o rețea de capilare foarte subțiri, prin pereții cărora sângele eliberează oxigen și este îmbogățit cu dioxid de carbon. Sângele venos se adună în vene și curge prin ele în atriu și din acesta în ventricul. În consecință, peștii au o singură circulație.
Sistemul circulator al amfibienilor este reprezentat de o inimă cu trei camere, formată din două atrii și un ventricul, și două cercuri de circulație a sângelui - mare (trunchi) și mic (pulmonar). Circulația pulmonară începe în ventricul, include vasele plămânilor și se termină în atriul stâng. Cercul cel mare începe și în ventricul. Sângele, după ce a trecut prin vasele întregului corp, se întoarce în atriul drept. Astfel, sângele arterial din plămâni intră în atriul stâng, iar sângele venos din întregul corp intră în atriul drept. Sângele arterial care curge din piele pătrunde și în atriul drept. Astfel, datorită aspectului circulației pulmonare, sângele arterial pătrunde și în inima amfibienilor. În ciuda faptului că sângele arterial și venos intră în ventricul, amestecarea completă a sângelui nu are loc din cauza prezenței buzunarelor și a partițiilor incomplete. Datorită lor, la părăsirea ventriculului, sângele arterial curge prin arterele carotide în cap, sângele venos în plămâni și piele și sângele amestecat în toate celelalte organe ale corpului. Astfel, la amfibieni nu există o separare completă a sângelui în ventricul, prin urmare intensitatea proceselor de viață este scăzută, iar temperatura corpului este variabilă.
Inima reptilelor este cu trei camere, dar amestecul complet de sânge arterial și venos nu are loc din cauza prezenței unui sept longitudinal incomplet. Trei vase care se extind din diferite părți ale ventriculului - artera pulmonară, arcurile aortice stângi și drepte - transportă sângele venos la plămâni, sângele arterial la cap și membrele anterioare și la părțile rămase - amestecat cu o predominanță a sângelui arterial. O astfel de alimentare cu sânge, precum și o capacitate scăzută de termoreglare, duce la faptul că
Temperatura corpului reptilelor depinde de condițiile de temperatură ale mediului.
Nivelul ridicat de activitate vitală a păsărilor se datorează unui sistem circulator mai avansat în comparație cu animalele din clasele anterioare. Au avut o separare completă a fluxurilor sanguine arteriale și venoase. Acest lucru se datorează faptului că inima păsărilor are patru camere și este complet împărțită în partea stângă - arterială și cea dreaptă - venoasă. Există un singur arc aortic (cel drept) și ia naștere din ventriculul stâng. Sângele arterial pur curge în el, furnizând toate țesuturile și organele corpului. Artera pulmonară pleacă din ventriculul drept, transportând sângele venos către plămâni. Sângele se mișcă rapid prin vase, schimbul de gaze are loc intens și se eliberează multă căldură. Sistemul circulator al mamiferelor nu este fundamental diferit de cel al păsărilor, spre deosebire de păsări, la mamifere arcul aortic stâng se îndepărtează de ventriculul stâng.
Întrebarea 22
DEZVOLTAREA ARCURILOR ARTERIALE
Arcurile arteriale, arcadele aortice, vasele de sânge care se formează la embrionii de vertebrate sub formă de 6-7 (în ciclostomi până la 15) trunchiuri laterale pereche care se extind din aorta abdominală. A. d. trec de-a lungul septurilor interbranhiale spre partea dorsală a faringelui și, unindu-se, formează aorta dorsală. Primele 2 perechi de arcade arteriale sunt de obicei reduse devreme; la pești și larvele de amfibieni se păstrează sub formă de vase mici. Restul de 4-5 perechi de arcade arteriale devin vase branchiale. La vertebratele terestre, arterele carotide se formează din a treia pereche de arcade arteriale, iar arterele pulmonare din a șasea. La amfibienii caudați, de obicei, a 4-a și a 5-a pereche de arcade arteriale formează trunchiurile sau rădăcinile aortei, fuzionând în aorta dorsală. La amfibienii și reptilele fără coadă, arcurile aortice iau naștere numai din a 4-a pereche de arcade arteriale, iar a 5-a este redusă. La păsări și mamifere, arcadele arteriale a 5-a și jumătate din a 4-a sunt reduse; la păsări, jumătatea dreaptă devine aortă; la mamifere, jumătatea stângă devine aortă. Uneori la indivizii adulți se păstrează vasele embrionare care leagă arcadele aortice cu arterele carotide (canalele carotide) sau pulmonare (canalele botaliene).
Întrebarea 23
Sistemul respirator.
Majoritatea animalelor sunt aerobe. Difuzia gazelor din atmosferă printr-o soluție apoasă are loc în timpul respirației. Elementele respirației pielii și apei sunt păstrate chiar și la vertebratele superioare. Pe parcursul evoluției, animalele au dezvoltat o varietate de aparate respiratorii - derivate ale pielii și tubului digestiv. Branhiile și plămânii sunt derivați ai faringelui.
FILGENEZA ORGANELOR RESPIRATORII
Organele respiratorii - branhiile - sunt situate pe partea superioară a celor patru arcade branhiale sub formă de petale de un roșu aprins. Apa intră în gura peștelui, este filtrată prin fantele branhiale, spălând branhiile și este evacuată de sub capacul branhiilor. Schimbul de gaze are loc în numeroase capilare branhiale, în care sângele curge spre apă spălând branhiile.
Broaștele respiră prin plămâni și prin piele. Plămânii sunt perechi de saci goale, cu o suprafață interioară celulară, pătrunși de o rețea de capilare sanguine, unde are loc schimbul de gaze. Mecanismul de respirație al amfibienilor este imperfect, de tip presiune. Animalul ia aer în cavitatea orofaringiană, pentru care coboară fundul gurii și deschide nările. Apoi nările se închid cu valve, podeaua gurii se ridică și aerul este forțat în plămâni. Aerul este eliminat din plămâni prin contractarea muşchilor pectorali. Suprafața plămânilor amfibienilor este mică, mai mică decât suprafața pielii.
Organe respiratorii - plămâni (reptile). Pereții lor au o structură celulară, ceea ce mărește semnificativ suprafața. Nu există respirație cutanată. Ventilația plămânilor este mai intensă decât la amfibieni și este asociată cu modificări ale volumului toracelui. Căile respiratorii - traheea, bronhiile - protejează plămânii de efectele de uscare și răcire ale aerului care vine din exterior.
Plămânii păsărilor sunt corpuri dense, spongioase. Bronhiile, care au intrat în plămâni, se ramifică puternic în ele până la cele mai subțiri, bronhiole închise orbește, încurcate într-o rețea de capilare, unde
iar schimbul de gaze are loc. Unele dintre bronhiile mari, fără ramificare, se extind dincolo de plămâni și se extind în saci de aer uriași cu pereți subțiri, al căror volum este de multe ori mai mare decât volumul plămânilor (Fig. 11.23). Sacii de aer sunt localizați între diferite organe interne, iar ramurile lor trec între mușchi, sub piele și în cavitățile oaselor.
Mamiferele respiră cu plămâni care au o structură alveolară, datorită căreia suprafața respiratorie depășește suprafața corpului de 50 de ori sau mai mult. Mecanismul de respirație este cauzat de o modificare a volumului toracelui datorită mișcării coastelor și a unui mușchi special caracteristic mamiferelor - diafragma.
Întrebarea 24
FILGENEZA CREIERULUI
Sistemul nervos central al peștilor este format din creier și măduva spinării. Creierul la pești, ca la toate vertebratele, este reprezentat de cinci secțiuni: creierul anterior, intermediar, mijlociu, cerebel și medular oblongata. Lobii olfactivi bine dezvoltați se extind din prosencefal. Mezencefalul, care analizează percepțiile vizuale, precum și cerebelul, care reglează coordonarea mișcărilor și menținerea echilibrului, realizează cea mai mare dezvoltare.
Creierul de amfibieni are aceleași cinci secțiuni ca și creierul de pește. Cu toate acestea, diferă de acesta prin dezvoltarea mai mare a creierului anterior, care la amfibieni este împărțit în două emisfere. Cerebelul este subdezvoltat din cauza mobilității reduse și monotoniei. modele diferite de mișcări ale amfibienilor.
Creierul reptilelor, în comparație cu cel al amfibienilor, are un cerebel mai bine dezvoltat și emisferele cerebrale ale prosencefalului, a căror suprafață are rudimentele cortexului. Acest lucru determină forme variate și mai complexe de comportament adaptativ.
Creierul păsărilor diferă de creierul reptilelor prin dimensiunea mare a emisferelor creierului anterior și cerebelului.
Creierul mamiferelor este relativ mare din cauza volumului crescut al creierului anterior și al cerebelului. Dezvoltarea creierului anterior are loc datorită creșterii acoperișului său - bolta cerebrală sau cortexul cerebral.
Întrebarea 25
FILGENEZA SISTEMELOR EXCRETOARE SI GENERALE
Organele excretoare ale peștilor sunt muguri perechi de trunchi în formă de panglică, localizați în cavitatea corpului sub coloana vertebrală. Au pierdut contactul cu cavitatea corpului și elimină deșeurile nocive, filtrăndu-le din sânge. La peștii de apă dulce, produsul final al metabolismului proteinelor este amoniacul toxic. Se dizolvă într-o cantitate mare de apă și, prin urmare, peștii excretă multă urină lichidă. Apa excretată în urină este ușor de completat datorită aportului constant prin piele, branhii și cu alimente. La peștii marini, produsul final al metabolismului azotului este ureea mai puțin toxică, a cărei eliminare necesită mai puțină apă. Urina formată în rinichi curge prin ureterele pereche în vezică, de unde este evacuată prin orificiul excretor. Glandele sexuale pereche - ovarele și testiculele - au canale excretoare. Fertilizarea la majoritatea peștilor este externă și are loc în apă.
Organele excretoare ale amfibienilor, ca si cele ale pestilor, sunt reprezentate de rinichii trunchiului. Cu toate acestea, spre deosebire de pești, aceștia au aspectul unor corpuri compacte turtite, întinse pe laterale
vertebra sacră. Rinichii contin glomeruli care filtreaza din sange produsele de degradare nocive (in principal uree) si in acelasi timp substante importante pentru organism (zaharuri, vitamine etc.). În timpul curgerii prin tubii renali, substanțele utile organismului sunt absorbite înapoi în sânge, iar urina curge prin două uretere în cloac și de acolo în vezică. După ce vezica urinară este umplută, pereții ei musculari se contractă, urina este eliberată în cloaca și aruncată afară. Pierderea de apă din corpul amfibienilor cu urină, precum și în pești, este completată prin aportul acesteia prin piele. Gonadele sunt împerecheate. Oviductele pereche se varsă în cloacă, iar canalul deferent în uretere.
Organele excretoare ale reptilelor sunt reprezentate de rinichii pelvieni, în care aria totală de filtrare a glomerulilor este mică, în timp ce lungimea tubilor este semnificativă. Aceasta promovează reabsorbția intensivă a apei filtrate de glomeruli în capilarele sanguine. În consecință, excreția de deșeuri la reptile are loc cu pierderi minime de apă. La acestea, ca și la artropodele terestre, produsul final al excreției este acidul uric, care necesită o cantitate mică de apă pentru a fi excretat din organism. Urina este colectată prin uretere în cloac și din aceasta în vezică, din care este excretată sub formă de suspensie de cristale mici.
Izolarea mamiferelor. Rinichii pelvieni ai mamiferelor sunt similari ca structura cu cei ai pasarilor. Urina cu un conținut ridicat de uree curge din rinichi prin uretere în vezică și din ea.
Întrebarea 26
Filogeneza tegumentului corporal:
Principalele direcții de evoluție a tegumentelor cordate:
1) diferențierea în două straturi: exterior - epidermă, interior - derm și o creștere a grosimii dermului;
1) de la epidermă cu un singur strat la multistrat;
2) diferențierea dermei în 2 straturi - papilar și reticular:
3) apariția grăsimii subcutanate și îmbunătățirea mecanismelor de termoreglare;
4) de la glande unicelulare la cele multicelulare;
5) diferențierea diverșilor derivați ai pielii.
În acordurile inferioare (lanceta) epiderma este monostratificată, cilindrică și are celule glandulare care secretă mucus. Dermul (coriumul) este reprezentat de un strat subțire de țesut conjunctiv neformat.
La vertebratele inferioare, epiderma devine multistratificată. Stratul său inferior este germinal (bazal), celulele sale se divid și reînnoiesc celulele straturilor de deasupra. Dermul are fibre, vase și nervi corect localizate.
Derivații pielii sunt: glandele mucoase unicelulare (la peștii ciclostom) și multicelulare (la amfibieni); solzi: a) placoid la pești cartilaginoși, la dezvoltarea cărora participă epiderma și derma; b) osos la peștii osoși, care se dezvoltă în detrimentul dermului.
Scama placoidă este acoperită la exterior cu un strat de smalț (origine ectodermică), sub care se află dentina și pulpa (origine mezodermă). Solzii și mucusul îndeplinesc o funcție de protecție.
Amfibienii au pielea subțire, netedă, fără solzi. Pielea conține un număr mare de glande mucoase multicelulare, a căror secreție hidratează tegumentul și are proprietăți bactericide. Pielea participă la schimbul de gaze.
La vertebratele superioare, datorită atingerii pământului, epiderma devine uscată și are strat cornos.
La reptile Se dezvoltă solzi excitați și glandele pielii sunt absente.
La mamifere: epiderma și derma sunt bine dezvoltate, aparețesut adipos subcutanat.
Întrebarea 27
FILGENEZA APARATULUI DIGESTIV.
Peștii mănâncă o varietate de alimente. Specializarea alimentară se reflectă în structura organelor digestive. Gura duce în cavitatea bucală, care conține de obicei numeroși dinți localizați pe maxilar, palatin și alte oase. Nu există glande salivare. Din cavitatea bucală, alimentele trec în faringe, perforate de fante branhiale, iar prin esofag intră în stomac, ale căror glande secretă abundent sucuri digestive. Unii pești (ciprinide și o serie de alții) nu au stomac și alimentele intră direct în intestinul subțire, unde, sub influența unui complex de enzime secretate de glandele intestinului însuși, ficatul și pancreasul, hrana este nutrienții dizolvați și dizolvați sunt absorbiți. Diferențierea sistemului digestiv al amfibienilor a rămas aproximativ la același nivel cu cel al strămoșilor lor - peștii. Cavitatea orofaringiană comună trece în esofagul scurt, în spatele căruia se află un stomac slab separat, care trece fără o limită ascuțită în intestin. Intestinul se termină cu rectul, care trece în cloaca. Canalele glandelor digestive - ficatul și pancreasul - curg în duoden. Conductele glandelor salivare, care sunt absente la pești, se deschid în cavitatea orofaringiană și udă cavitatea bucală și alimentele. Stilul de viață terestru este asociat cu apariția unei adevărate limbi în cavitatea bucală, principalul organ de obținere a hranei.
În sistemul digestiv al reptilelor, diferențierea în secțiuni este mai bună decât cea a amfibienilor. Hrana este captată de fălci care au dinți pentru a ține prada. Cavitatea bucală este mai bine delimitată de faringe decât la amfibieni. În partea inferioară a cavităţii bucale se află o limbă mobilă, bifurcată la capăt. Mâncarea este umezită cu salivă, ceea ce face mai ușor de înghițit. Esofagul este lung din cauza dezvoltării gâtului. Stomacul, delimitat de esofag, are pereți musculari. La marginea intestinului subțire și gros există un cecum. Canalele hepatice și pancreasului
Glandele se deschid în duoden. Timpul necesar pentru digerarea alimentelor depinde de temperatura corpului reptilelor.
Sistemul digestiv al mamiferelor. Dinții se află în celulele oaselor maxilarului și sunt împărțiți în incisivi, canini și molari. Deschiderea gurii este înconjurată de buze cărnoase, ceea ce este caracteristic doar mamiferelor datorită hrănirii cu lapte. În cavitatea bucală, alimentele, pe lângă faptul că sunt mestecate de dinți, sunt afectate chimic de enzimele salivare, iar apoi trec succesiv în esofag și stomac. Stomacul la mamifere este bine separat de alte părți ale tractului digestiv și este echipat cu glande digestive. La majoritatea speciilor de mamifere, stomacul este împărțit în mai multe sau mai puține secțiuni. Este cel mai complex la artiodactilii rumegătoare. Intestinul are secțiuni subțiri și groase. La marginea secțiunilor subțiri și groase iese cecumul, în care se fermentează fibra. Canalele ficatului și pancreasului se deschid în cavitatea duodenului.
Întrebarea 28
Sistemul endocrin.
Orice organism produce compuși care sunt distribuiți în tot organismul și au rol integrator. Plantele au fitohormoni care controlează creșterea, dezvoltarea fructelor, florilor, dezvoltarea mugurilor axilari, diviziunea cambiului etc. Algele unicelulare au fitohormoni.
Hormonii au apărut în organismele multicelulare când au apărut celule endocrine speciale. Cu toate acestea, compușii chimici care joacă rolul hormonilor au existat înainte. Tiroxina, triiodotironina (glanda tiroida) au fost gasite in cianobacterii. Reglarea hormonală la insecte este puțin înțeleasă.
În 1965, Wilson a izolat insulina din stele de mare.
S-a dovedit că definirea hormonului este foarte dificilă.
Hormonul este o substanță chimică specifică secretată de anumite celule într-o anumită zonă a corpului, care intră în fluxul sanguin și apoi are un efect specific asupra anumitor celule sau organe țintă situate în alte zone ale corpului, ceea ce duce la coordonarea funcțiile întregului organism ca întreg.
Se cunosc un număr mare de hormoni de mamifere. Ele sunt împărțite în 3 grupuri principale.
Feromonii. Eliberat în mediul extern. Cu ajutorul lor, animalele primesc și transmit informații. La om, mirosul acidului 14-hidroxitetradecanoic se distinge clar doar de femeile care au ajuns la pubertate.
Cele mai simplu organizate organisme multicelulare - de exemplu, bureții au, de asemenea, o aparență de sistem endocrin. Bureții sunt formați din 2 straturi - endodermul și exodermul; între ele se află mezenchimul, care conține compuși macromoleculari caracteristici țesutului conjunctiv al organismelor mai bine organizate. Mezenchimul conține celule migratoare; unele celule sunt capabile să secrete serotonină și acetilcolină. Bureții nu au sistem nervos. Substanțele sintetizate în mezenchim servesc la conectarea părților individuale ale corpului. Coordonarea are loc prin mișcarea celulelor de-a lungul mezenchimului. Există și transferul de substanțe între celule. Se pune bazele semnalizării chimice, ceea ce este caracteristic altor animale. Nu există celule endocrine independente.
Celenteratele au un sistem nervos primitiv. Inițial, celulele nervoase au îndeplinit o funcție neurosecretoare. Funcția trofică a controlat creșterea și dezvoltarea organismului. Apoi celulele nervoase au început să se întindă și să formeze procese lungi. Secreția a fost eliberată în apropierea organului țintă, fără transfer (din moment ce nu era sânge). Mecanismul endocrin a apărut mai devreme decât mecanismul de conducere. Celulele nervoase au fost endocrine, iar apoi au dobândit proprietăți conducătoare. Celulele neurosecretoare au fost primele celule secretoare.
Protostomii și deuterostomii produc aceiași hormoni steroizi și peptidici. Este general acceptat că în procesul de evoluție pot apărea altele noi (mutații, duplicări de gene) din unii hormoni polipeptidici. Dublările sunt mai puțin suprimate de selecția naturală decât mutațiile. Mulți hormoni pot fi sintetizați nu într-o singură glandă, ci în mai multe. De exemplu, insulina este produsă în pancreas, glanda submandibulară, duoden și alte organe. Există o dependență a genelor care controlează sinteza hormonală de poziție.
