Регенерация как свойство живого: способность к самообновлению и восстановлению. Типы регенерации
РЕГЕНЕРАЦИЯ
восстановление организмом утраченных частей на той или иной стадии жизненного цикла. Регенерация обычно происходит в случае повреждения или утраты какого-нибудь органа или части организма. Однако помимо этого в каждом организме на протяжении всей его жизни постоянно идут процессы восстановления и обновления. У человека, например, постоянно обновляется наружный слой кожи. Птицы периодически сбрасывают перья и отращивают новые, а млекопитающие сменяют шерстный покров. У листопадных деревьев листья ежегодно опадают и заменяются свежими. Такую регенерацию, обычно не связанную с повреждениями или утратой, называют физиологической. Регенерацию, происходящую после повреждения или утраты какой-либо части тела, называют репаративной. Здесь мы рассмотрим только репаративную регенерацию. Репаративная регенерация может быть типичной или атипичной. При типичной регенерации утраченная часть замещается путем развития точно такой же части. Причиной утраты может быть внешнее воздействие (например, ампутация), или же животное намеренно отрывает часть своего тела (аутотомия), как ящерица, обламывающая часть своего хвоста, спасаясь от врага. При атипичной регенерации утраченная часть замещается структурой, отличающейся от первоначальной количественно или качественно. У регенерировавшей конечности головастика число пальцев может оказаться меньше исходного, а у креветки вместо ампутированного глаза может вырасти антенна.
РЕГЕНЕРАЦИЯ У ЖИВОТНЫХ
Способность к регенерации широко распространена среди животных. Вообще говоря, низшие животные чаще способны к регенерации, чем более сложные высокоорганизованные формы. Так, среди беспозвоночных гораздо больше видов, способных восстанавливать утраченные органы, чем среди позвоночных, но только у некоторых из них возможна регенерация целой особи из небольшого ее фрагмента. Тем не менее общее правило о снижении способности к регенерации с повышением сложности организма нельзя считать абсолютным. Такие примитивные животные, как гребневики и коловратки, практически не способны к регенерации, а у гораздо более сложных ракообразных и амфибий эта способность хорошо выражена; известны и другие исключения.
Некоторые близкородственные животные сильно различаются в этом отношении. Так, у дождевого червя из небольшого кусочка тела может полностью регенерировать новая особь, тогда как пиявки неспособны восстановить один утраченный орган. У хвостатых амфибий на месте ампутированной конечности образуется новая, а у лягушки культя просто заживает и никакого нового роста не происходит. Многие беспозвоночные способны к регенерации значительной части тела. У губок, гидроидных полипов, плоских, ленточных и кольчатых червей, мшанок, иглокожих и оболочников из небольшого фрагмента тела может регенерировать целый организм. Особенно примечательна способность к регенерации у губок. Если тело взрослой губки продавить через сетчатую ткань, то все клетки отделятся друг от друга, как просеянные сквозь сито. Если затем поместить все эти отдельные клетки в воду и осторожно, тщательно перемешать, полностью разрушив все связи между ними, то спустя некоторое время они начинают постепенно сближаться и воссоединяются, образуя целую губку, сходную с прежней. В этом участвует своего рода "узнавание" на клеточном уровне, о чем свидетельствует следующий эксперимент. Губки трех разных видов разделяли описанным способом на отдельные клетки и как следует перемешивали. При этом обнаружилось, что клетки каждого вида способны "узнавать" в общей массе клетки своего вида и воссоединяются только с ними, так что в результате образовалась не одна, а три новых губки, подобные трем исходным.
Ленточный червь, длина которого во много раз превышает его ширину, способен воссоздать целую особь из любого участка своего тела. Теоретически возможно, разрезав одного червя на 200 000 кусочков, получить из него в результате регенерации 200 000 новых червей. Из одного луча морской звезды может регенерировать целая звезда.

Моллюски, членистоногие и позвоночные не способны регенерировать целую особь из одного фрагмента, однако у многих из них происходит восстановление утраченного органа. Некоторые в случае необходимости прибегают к аутотомии. Птицы и млекопитающие как эволюционно наиболее продвинутые животные меньше других способны к регенерации. У птиц возможно замещение перьев и некоторых частей клюва. Млекопитающие могут восстанавливать покров, когти и частично печень; они способны также к заживлению ран, а олени - к отращиванию новых рогов взамен сброшенных.
Процессы регенерации. В регенерации у животных участвуют два процесса: эпиморфоз и морфаллаксис. При эпиморфической регенерации утраченная часть тела восстанавливается за счет активности недифференцированных клеток. Эти клетки, похожие на эмбриональные, накапливаются под пораненным эпидермисом у поверхности разреза, где они образуют зачаток, или бластему. Клетки бластемы постепенно размножаются и превращаются в ткани нового органа или части тела. При морфаллаксисе другие ткани тела или органа непосредственно преобразуются в структуры недостающей части. У гидроидных полипов регенерация происходит главным образом путем морфаллаксиса, а у планарий в ней одновременно участвуют и эпиморфоз, и морфаллаксис. Регенерация путем образования бластемы широко распространена у беспозвоночных и играет особенно важную роль в регенерации органов у амфибий. Существует две теории происхождения бластемных клеток: 1) клетки бластемы происходят из "резервных клеток", т.е. клеток, оставшихся неиспользованными в процессе эмбрионального развития и распределившихся по разным органам тела; 2) ткани, целостность которых была нарушена при ампутации, "дедифференцируются" в области разреза, т.е. дезинтегрируются и превращаются в отдельные бластемные клетки. Таким образом, согласно теории "резервных клеток", бластема образуется из клеток, остававшихся эмбриональными, которые мигрируют из разных участков тела и скапливаются у поверхности разреза, а согласно теории "дедифференцированной ткани", бластемные клетки происходят из клеток поврежденных тканей. В подтверждение как одной, так и другой теории имеется достаточно данных. Например, у планарий резервные клетки более чувствительны к рентгеновским лучам, чем клетки дифференцированной ткани; поэтому их можно разрушить, строго дозируя облучение, чтобы не повредить нормальные ткани планарии. Облученные таким образом особи выживают, но утрачивают способность к регенерации. Однако если только переднюю половину тела планарии подвергнуть облучению, а затем разрезать, то регенерация происходит, хотя и с некоторой задержкой. Задержка свидетельствует о том, что бластема образуется из резервных клеток, мигрирующих на поверхность разреза из необлученной половины тела. Миграцию этих резервных клеток по облученной части тела можно наблюдать под микроскопом. Сходные эксперименты показали, что у тритона регенерация конечностей происходит за счет бластемных клеток местного происхождения, т.е. за счет дедифференцировки поврежденных тканей культи. Если, например, облучить всю личинку тритона, за исключением, скажем, правой передней конечности, а затем ампутировать эту конечность на уровне предплечья, то у животного отрастает новая передняя конечность. Очевидно, что необходимые для этого бластемные клетки поступают именно из культи передней конечности, так как все остальное тело подверглось облучению. Более того, регенерация происходит даже в том случае, если облучают всю личинку, за исключением участка шириной 1 мм на правой передней лапке, а затем последнюю ампутируют, производя разрез через этот необлученный участок. В этом случае совершенно очевидно, что бластемные клетки поступают с поверхности разреза, поскольку все тело, включая правую переднюю лапку, было лишено способности к регенерации. Описанные процессы анализировали с применением современных методов. Электронный микроскоп позволяет наблюдать изменения в поврежденных и регенерирующих тканях во всех деталях. Созданы красители, выявляющие определенные химические вещества, содержащиеся в клетках и тканях. Гистохимические методы (с применением красителей) дают возможность судить о биохимических процессах, происходящих при регенерации органов и тканей.
Полярность. Одна из самых загадочных проблем в биологии - происхождение полярности у организмов. Из шаровидного яйца лягушки развивается головастик, у которого с самого начала на одном конце тела находится голова с головным мозгом, глазами и ртом, а на другом - хвост. Подобным же образом, если разрезать тело планарии на отдельные фрагменты, на одном конце каждого фрагмента развивается голова, а на другой - хвост. При этом голова всегда образуется на переднем конце фрагмента. Эксперименты ясно показывают, что у планарии существует градиент метаболической (биохимической) активности, проходящий по передне-задней оси ее тела; при этом наивысшей активностью обладает самый передний конец тела, а в направлении к заднему концу активность постепенно снижается. У любого животного голова всегда образуется на том конце фрагмента, где метаболическая активность выше. Если направление градиента метаболической активности в изолированном фрагменте планарии изменить на противоположное, то и формирование головы произойдет на противоположном конце фрагмента. Градиент метаболической активности в теле планарий отражает существование какого-то более важного физико-химического градиента, природа которого пока неизвестна. В регенерирующей конечности тритона полярность новообразуемой структуры, по-видимому, определяется сохранившейся культей. По причинам, которые еще остаются неясными, в регенерирующем органе формируются только структуры, расположенные дистальнее раневой поверхности, а те, что расположены проксимальнее (ближе к телу), не регенерируют никогда. Так, если ампутировать кисть тритона, а оставшуюся часть передней конечности вставить обрезанным концом в стенку тела и дать этому дистальному (отдаленному от тела) концу прижиться на новом, необычном для него месте, то последующая перерезка этой верхней конечности вблизи плеча (освобождающая ее от связи с плечом) приводит к регенерации конечности с полным набором дистальных структур. У такой конечности имеются на момент перерезки следующие части (начиная с запястья, слившегося со стенкой тела): запястье, предплечье, локоть и дистальная половина плеча; затем, в результате регенерации, появляются: еще одна дистальная половина плеча, локоть, предплечье, запястье и кисть. Таким образом, инвертированная (перевернутая) конечность регенерировала все части, расположенные дистальнее раневой поверхности. Это поразительное явление указывает на то, что ткани культи (в данном случае культи конечности) контролируют регенерацию органа. Задача дальнейших исследований - выяснить, какие именно факторы контролируют этот процесс, что стимулирует регенерацию и что заставляет клетки, обеспечивающие регенерацию, скапливаться на раневой поверхности. Некоторые ученые полагают, что поврежденные ткани выделяют какой-то химический "раневой фактор". Однако выделить химическое вещество, специфичное для ран, пока не удалось.
РЕГЕНЕРАЦИЯ У РАСТЕНИЙ
Широкое распространение регенерации в царстве растений обусловлено сохранением у них меристем (тканей, состоящих из делящихся клеток) и недифференцированных тканей. В большинстве случаев регенерация у растений - это, в сущности, одна из форм вегетативного размножения. Так, на кончике нормального стебля имеется верхушечная почка, обеспечивающая непрерывное образование новых листьев и рост стебля в длину в течение всей жизни данного растения. Если отрезать эту почку и поддерживать ее во влажном состоянии, то из имеющихся в ней паренхимных клеток или из каллуса, образующегося на поверхности среза, часто развиваются новые корни; почка при этом продолжает расти и дает начало новому растению. То же самое происходит в природе, когда отламывается ветка. Плети и столоны разделяются в результате отмирания старых участков (междоузлий). Таким же образом разделяются корневища ириса, волчьей стопы или папоротников, образуя новые растения. Обычно клубни, например клубни картофеля, продолжают жить после отмирания подземного стебля, на котором они выросли; с наступлением нового вегетационного периода они могут дать начало собственным корням и побегам. У луковичных растений, например у гиацинтов или тюльпанов, побеги формируются у основания чешуй луковицы и могут в свою очередь образовывать новые луковицы, которые в конечном счете дают корни и цветоносные стебли, т.е. становятся самостоятельными растениями. У некоторых лилейных воздушные луковички образуются в пазухах листьев, а у ряда папоротников на листьях вырастают выводковые почки; в какой-то момент они опадают на землю и возобновляют рост. Корни менее способны к образованию новых частей, чем стебли. Клубню георгина для этого необходима почка, образующаяся у основания стебля; однако батат может дать начало новому растению из почки, образуемой корневой шишкой. Листья тоже способны к регенерации. У некоторых видов папоротников, например у кривокучника (Camptosorus), листья сильно вытянуты и имеют вид длинных волосовидных образований, заканчивающихся меристемой. Из этой меристемы развивается зародыш с зачаточными стеблем, корнями и листьями; если кончик листа родительского растения наклонится вниз и соприкоснется с землей или мхом, зачаток начинает расти. Новое растение отделяется от родительского после истощения этого волосовидного образования. Листья суккулентного комнатного растения каланхое несут по краям хорошо развитые растеньица, которые легко отпадают. Новые побеги и корни формируются на поверхности листьев бегонии. Специальные тельца, называемые зародышевыми почками, развиваются на листьях некоторых плауновых (Lycopodium) и печеночников (Marchantia); упав на землю, они укореняются и образуют новые зрелые растения. Многие водоросли успешно размножаются, расчленяясь на фрагменты под ударами волн.
См. также СИСТЕМАТИКА РАСТЕНИЙ . ЛИТЕРАТУРА Мэттсон П. Регенерация - настоящее и будущее. М., 1982 Гилберт С. Биология развития, тт. 1-3. М., 1993-1995
Энциклопедия Кольера. - Открытое общество . 2000 .
Синонимы :Смотреть что такое "РЕГЕНЕРАЦИЯ" в других словарях:
РЕГЕНЕРАЦИЯ - РЕГЕНЕРАЦИЯ, процесс образования нового, органа или ткани на месте удаленного тем или иным образом участка организма. Очень часто Р. определяется как процесс восстановления утраченного, т.. е. образование органа, подобного удаленному. Такое… … Большая медицинская энциклопедия
- (поздн. лат., от лат. re опять, вновь, и genus, eris род, поколение). Возрождение, возобновление, восстановление того, что было разрушено. В фигуральном значении: перемена к лучшему. Словарь иностранных слов, вошедших в состав русского языка.… … Словарь иностранных слов русского языка
РЕГЕНЕРАЦИЯ, в биологии способность организма к замещению одной из утраченных частей. Термин регенерация также относится к форме БЕСПОЛОГО РАЗМНОЖЕНИЯ, при котором новая особь возникает из отделенной части материнского организма … Научно-технический энциклопедический словарь
Восстановление, рекуперация; возмещение, регенерирование, возобновление, гетероморфоз, петтенкоферирование, возрождение, морфаллаксис Словарь русских синонимов. регенерация сущ., кол во синонимов: 11 возмещение (20) … Словарь синонимов
1) восстановление с помощью определенных физико химических процессов исходных состава и свойств отработанных продуктов для повторного их использования. В военном деле широкое распространение получила регенерация воздуха (особенно на подводных… … Морской словарь
Регенерация - – возвращение отработанному продукту исходных свойств. [Терминологический словарь по бетону и железобетону. ФГУП «НИЦ «Строительство» НИИЖБ им. А. А. Гвоздева, Москва, 2007 г. 110 стр.] Регенерация – восстановление отработанных… … Энциклопедия терминов, определений и пояснений строительных материалов
РЕГЕНЕРАЦИЯ - (1) восстановление исходных свойств и состава отработавших материалов (воды, воздуха, масел, резины и др.) для их повторного использования. Осуществляется с помощью определённых физ. хим. процессов в специальных устройствах регенераторах. Широко… … Большая политехническая энциклопедия
- (от позднелат. regeneratio возрождение возобновление), в биологии восстановление организмом утраченных или поврежденных органов и тканей, а также восстановление целого организма из его части. В большей степени присуща растениям и беспозвоночным… …
В технике,1) возвращение отработавшему продукту исходных качеств, напр. восстановление свойств отработавшей формовочной смеси в литейном производстве, очистка отработавшего смазочного масла, превращение изношенных резиновых изделий в пластичную… … Большой Энциклопедический словарь
РЕГЕНЕРАЦИЯ, регенерации, мн. нет, жен. (лат. regeneratio восстановление, возвращение). 1. Нагрев газа и воздуха, поступающих в печь, отработанными продуктами горения (тех.). 2. Воспроизведение животными утерянных органов (зоол.). 3. Излучение… … Толковый словарь Ушакова
Регенерация (от лат. regeneratio — возрождение) — процесс восстановления биологических структур в ходе жизнедеятельности организма. Регенерация поддерживает строение и функции организма, его целостность.Регенерационные процессы реализуются на разных уровнях организации — молекулярно-генетическом, субклеточном, клеточном, тканевом, органном, организменном.На молекулярно-генетическом уровне осуществляется репликация ДНК, ее репарация, синтез новых ферментов, молекул АТФ и т.д. Все эти процессы входят в обмен веществ клетки.На субклеточном уровне происходит восстановление структур клетки за счет образования новых структурных единиц и сборки органелл или деления сохранившихся органелл. Например, подвижные образования клеточной мембраны — рецепторы, ионные каналы и насосы — могут перемещаться, концентрироваться или распределяться в составе мембраны. Помимо этого они выходят из мембраны, разрушаются и заменяются новыми. Так, в миобластах каждую минуту деградирует и заменяется новыми молекулами примерно 1 мкм2 поверхности. В фоторецептор-ных клетках — палочках (рис. 8.73) есть наружный сегмент, состоящий примерно из тысячи так называемых фоторецепторных дисков — плотно уложенных участков клеточной мембраны, в которые погружены светочувствительные белки, связанные со зрительным пигментом. Эти диски непрерывно обновляются — деградируют на наружном конце и вновь возникают на внутреннем со скоростью 3-4 диска в час. Аналогично осуществляются процессы восстановления после повреждений. Воздействие митохондриальных ядов вызывает утрату крист митохондрий. После прекращения действия яда в печеночной клетке митохондрии восстанавливают свою структуру за 2-3 сут.Клеточный уровень регенерации подразумевает восстановление структуры и, в некоторых случаях, функций клетки. К примерам такого рода относится восстановление отростка нервной клетки нейрона. У млекопитающих этот процесс идет со скоростью 1 мм в сутки. Восстановление функций клетки может осуществляться за счет гиперплазии — увеличения количества внутриклеточных органелл (внутриклеточная регенерация).На следующем уровне — тканевом или клеточно-популяционном — происходит восполнение теряемых клеток определенного направления дифференцировки. Происходят перестройки в пределах клеточных по-пуляций, и их результатом становится восстановление функций ткани. Так, у человека время жизни клеток кишечного эпителия — 4-5 сут, тромбоцитов — 5-7 сут, эритроцитов — 120-125 сут. Ежесекундно разрушается порядка 1 млн эритроцитов и столько же образуется в красном костном мозге вновь. Возможность восстановления утраченных клеток обеспечивается благодаря тому, что в тканях существует два клеточных компартмента. Один — дифференцированные рабочие клетки, а другой — камбиальные клетки, способные к делению и последующей дифференцировке. Эти последние в настоящее время называют региональными стволовыми клетками (см. пп. 3.1.2, 3.2). Они коммити-рованы, т.е. судьба их предопределена (см. п. 8.3.1), поэтому они способны дать начало одному или нескольким определенным клеточным типам. Их дальнейшая дифференцировка определяется сигналами, поступающими извне: от окружения (межклеточными взаимодействиями) и дистантными (например, гормонами), в зависимости от которых в клетках избирательно активируются конкретные гены. Так, в эпителии тонкой кишки камбиальные клетки находятся в придонных зонах крипт (рис. 8.74). При определенных воздействиях они способны дать начало клеткам «каемчатого» всасывающего эпителия и некоторым одноклеточным железам.Органный уровень регенерации предполагает восстановление функции или структуры органа. На этом уровне наблюдаются не только преобразования клеточных популяций, но также и морфогенетические процессы. При этом реализуются те же механизмы, что и при формировании органов в эмбриогенезе. Та- Рис. 8.73. Схематическое изображение фоторецептора сетчатки — палочки: 1 — синаптическое тельце, примыкающее к нейральному слою сетчатки, 2 — ядро, 3 — аппарат Гольджи, 4 — внутренний сегмент с митохондриями, 5 — соединительная ресничка, 6 — наружный сегмент с фото-рецепторными дискамикая регенерация может осуществляться путем эпиморфоза, морфолаксиса, регенерационной гипертрофии. Эти способы и механизмы регенерации обсуждаются далее. На организменном уровне возможно в отдельных случаях воссоздание целостного организма из одной или группы клеток. Различают два вида регенерации: физиологическую и репаративную. Физиологическая (гомеостати-ческая) регенерация представляет собой процесс восстановления структур, которые снашиваются в процессе нормальной жизнедеятельности. Благодаря ей поддерживается структурный гомеостаз и обеспечивается возможность постоянного выполнения органами их функций. С общебиологической точки зрения физиологическая регенерация, как и обмен веществ, является проявлением такого важнейшего свойства жизни, как самообновление. Самообновление обеспечивает существование организма во времени и пространстве. В его основе лежит биогенная миграция атомов. На внутриклеточном уровне значение физиологической регенерации особенно велико для так называемых «вечных» тканей, утративших способность к регенерации путем деления клеток. В первую очередь это относится к нервной ткани, сетчатке глаза. На клеточном и тканевом уровнях осуществляется физиологическая регенерация в «лабильных» тканях, где Рис. 8.74. Локализация региональных стволовых клеток в эпителии тонкой кишки: 1 — неде-лящиеся клетки; 2 — делящиеся стволовые клетки; 3 — быстро делящиеся клетки; 4 — неделящиеся дифференцированные клетки; 5 — направление перемещения клеток; 6 — клетки, слущенные с поверхности кишечной ворсиныинтенсивность клеточного обновления очень велика, и в «растущих» тканях, клетки которых обновляются значительно медленнее. К первой группе относятся, например, роговица глаза, эпителий слизистой оболочки кишечника, клетки периферической крови, эпидермис кожи и его производные — волосы и ногти. Клетки таких органов, как печень, почка, надпочечник составляют вторую из указанных групп.Об интенсивности пролиферации судят по числу митозов, приходящихся на 1000 подсчитанных клеток. Если учесть, что сам митоз в среднем длится около 1 ч, а весь митотический цикл в соматических клетках в среднем протекает 22-24 ч, становится ясно, что для определения интенсивности обновления клеточного состава тканей необходимо подсчитать число митозов в течение одних или нескольких суток. Оказалось, что число делящихся клеток не одинаково в разные часы суток. Так был открыт суточный ритм клеточных делений, пример которого изображен на рис. 8.75.Суточный ритм числа митозов обнаружен не только в нормальных, но и в опухолевых тканях. Он отражает более общую закономерность, Рис. 8.75. Суточные изменения митотиче-ского индекса (МИ) в эпителии пищевода (1) и роговицы (2) мышей. Митотический индекс выражен в промилле (0/00), отражающем число митозов в тысяче подсчитанных клетока именно, ритмичность всех функций организма. Одна из современных областей биологии — хронобиология — изучает, в частности, механизмы регуляции суточных ритмов митотической активности, что имеет весьма большое значение для медицины. Существование самой суточной периодичности числа митозов указывает на регулируемость физиологической регенерации организмом. Кроме суточных, существуют лунные и годичные циклы обновления тканей и органов. Физиологическая регенерация присуща организмам всех видов, но особенно интенсивно она протекает у теплокровных позвоночных, так как у них вообще очень высока интенсивность функционирования всех органов по сравнению с другими животными. Репаративная регенерация (от лат. reparatio — восстановление) — восстановление биологических структур после травм и действия других повреждающих факторов. К таким факторам могут быть отнесены ядовитые вещества, болезнетворные агенты, высокие и низкие температуры (ожоги и обморожения), лучевые воздействия, голодание и т.д. Способность к регенерации не имеет однозначной зависимости от уровня организации, хотя давно уже было замечено, что более низко организованные животные обладают лучшей способностью к регенерации наружных органов. Это подтверждается удивительными примерами регенерации гидры, планарий, кольчатых червей, членистоногих, иглокожих, низших хордовых, например асцидий. Из позвоночных наилучшей регенерационной способностью обладают хвостатые земноводные. Известно, что разные виды одного и того же класса могут сильно отличаться по способности к регенерации. Кроме того, при изучении способности к регенерации внутренних органов оказалось, что она значительно выше у теплокровных животных, например у млекопитающих, по сравнению с земноводными. Регенерация у млекопитающих отличается своеобразием. Для регенерации некоторых наружных органов нужны особые условия. Язык, ухо, например, не регенерируют при краевом повреждении (фактически речь идет об ампутации краевой части структуры). Если же нанести сквозной дефект через всю толщу органа, восстановление идет хорошо. Регенерация внутренних органов может идти очень активно. Из небольшого фрагмента яичника восстанавливается целый орган. Есть предположение, что невозможность регенерации конечностей и других наружных органов у млекопитающих носит приспособительный характер и обусловлена отбором, поскольку при активном образе жизни требующие сложной регуляции морфогенетические процессы затрудняли бы существование. Ряд исследователей полагает, что организмы первоначально имели два способа исцеления от ран — действие иммунной системы и регенерацию. Но в ходе эволюции они стали несовместимы друг с другом. Хотя регенерация может показаться лучшим выбором, для нас более важны Т-клетки иммунной системы — основное оружие против опухолей. Регенерация конечности становится бессмысленной, если одновременно в организме бурно развиваются раковые клетки. Получается, что иммунная система, защищая нас от инфекций и рака, одновременно подавляет наши способности к восстановлению.Объем репаративной регенерации может быть очень разным.Крайний вариант — восстановление целого организма из отдельной малой его части, фактически из группы соматических клеток. Среди животных такое восстановление возможно у губок и кишечнополостных. Регенерацию гидры можно осуществить из группы клеток, полученных при продавливании ее через сито. Среди растений возможно развитие целого нового растения даже из одной соматической клетки, как это получено на примере моркови и табака. Такой вид восстановительных процессов сопровождается возникновением новой морфогенетической оси организма и назван Б.П. Токиным «соматическим эмбриогенезом», так как во многом напоминает эмбриональное развитие. В качестве подобного варианта регенерации может рассматриваться клонирование в эксперименте целого организма из одной соматической клетки у млекопитающих.Следующий по объему вариант — восстановление больших участков организма, состоящих из комплекса органов. Примером служит регенерация у гидры, ресничного червя (планарии), морской звезды (рис. 8.76). При удалении части животного из оставшегося фрагмента, даже очень небольшого, возможно восстановление полноценного организма. Например, восстановление морской звезды из сохранившегося луча.Далее в этом ряду следует регенерация отдельных органов, которая широко распространена в животном царстве, например, хвоста у ящерицы, глаз у членистоногих, глаза, конечности, хвоста у тритона.Заживление кожных покровов, ран, повреждений костей и других внутренних органов — наименее объемный процесс, но не менее важный для восстановления структурно-функциональной целостности организма.Существует несколько способов репаративной регенерации. К ним относят эпиморфоз, морфаллаксис, регенерационную гипертрофию, компенсаторную гипертрофию, заживление эпителиальных ран, тканевую регенерацию. Рис. 8.76. Регенерация комплекса органов у некоторых видов беспозвоночных животных: а — гидра; б — плоский червь; в — морская звезда; г — восстановление морской звезды из лучаЭпиморфоз представляет собой наиболее очевидный способ регенерации, заключающийся в отрастании нового органа от ампутационной поверхности. Иллюстрацией может служить регенерация хрусталика или конечности у хвостатых амфибий (рис. 8.77). Рассмотрим более детально процесс регенерации на примере эпиморфоза конечности тритона. В процессе восстановления выделяют регрессивную и прогрессивную фазы регенерации. Регрессивная фаза начинается с заживления раны, во время которого происходят следующие основные события: остановка Рис. 8.77. Регенерация хрусталика (1) из дорзальной радужки (2) у тритонакровотечения, сокращение мягких тканей культи конечности, образование над раневой поверхностью сгустка фибрина и миграция эпидермиса, покрывающего ампутационную поверхность.Затем начинается разрушение тканей непосредственно проксималь-нее места ампутации. Одновременно в разрушенные мягкие ткани проникают клетки, участвующие в воспалительном процессе, наблюдается фагоцитоз и местный отек. Вслед за этим в области под раневым эпидермисом начинается дедифференцировка специализированных клеток: мышечных, костных, хрящевых и т.д. Клетки приобретают черты мезенхимных, образуют скопление и формируют регенерационную бластему (рис. 8.78). В это же время раневой эпидермис быстро утолщается и образует апикальную эктодермальную шапочку. На этом этапе в регенерационную бластему и эктодермальную шапочку врастают сосуды и нервные волокна.Далее начинается прогрессивная фаза, для которой наиболее характерны процессы роста и морфогенеза. Длина и масса регенерационной бластемы быстро увеличиваются. Она приобретает коническую форму. Мезенхимные клетки бластемы дедифференцируются, давая начало всем специализированным клеточным типам, которые необходимы для формирования структур конечности. Осуществляется рост конечности и ее морфогенез (формообразование). Когда форма конечности в общих чертах уже сложилась, регенерат все еще меньше нормальной конечности. Чем крупнее животное, тем больше эта разница в размерах. Для завершения морфогенеза требуется время, по истечении которого регенерат достигает размеров нормальной конечности.Некоторые стадии восстановления передней конечности у тритона после ампутации на уровне плеча показаны на рис. 8.79. Рис. 8.78. Регенерация конечности у тритона: а — нормальная конечность, б — ампутация; в — формирование апикальной шапочки и бластемы; г — редиф-ференцировка клеток; д — вновь сформированная конечность. 1 — бластема; 2 — апикальная эктодермальная шапочка; 3 — редифференцировка клеток бластемы (пояснения в тексте)У молодых личинок аксолотлей конечность может регенерировать за 3 нед, у взрослых тритонов и аксолотлей — за 1-2 мес, а у наземных амбистом для этого требуется около 1 года.Морфаллаксис — регенерация путем перестройки регенерирующего участка. Примером служит регенерация гидры из кольца, вырезанного из середины ее тела, или восстановление планарии из одной десятой или двадцатой ее части. На раневой поверхности в этом случае не происходит значительных формообразовательных процессов. Отрезанный кусочек сжимается, клетки внутри него перестраиваются, и возникает целая особь уменьшенных размеров, которая затем растет. Этот способ регенерации впервые описал Т. Морган в 1900 г. В соответствии с его описанием, морфаллаксис осуществляется без митозов. Нередко имеет место сочетание эпиморфного роста наместе ампутации с реорганизацией путем морфаллаксиса в прилежащих частях тела.Регенерационная гипертрофия (эндоморфоз) относится к внутренним органам. Этот способ регенерации заключается в увеличении размеров остатка органа без восстановления исходной формы. Иллюстрацией служит регенерация печени позвоночных, в том числе млекопитающих. При краевом ранении печени удаленная часть органа никогда не восстанавливается. Раневая поверхность заживает. В то же время вну- Рис. 8.79. Регенерация передней конечности у тритона в эксперименте Рис. 8.80. Влияние возраста на увеличение числа клубочков нефронов после удаления одной почки у крыс вскоре после рождения: 1 — кривая прироста числа клубочков в нормальном постнатальном развитии в одной почке; 2 — кривые увеличения числа вновь образуемых клубочков после удаления почки на разных сроках онтогенезатри оставшейся части усиливается размножение клеток (гиперплазия) и даже после удаления 2/3 печени восстанавливаются исходные масса и объем, но не форма. Внутренняя структура печени оказывается нормальной, дольки имеют типичную для них величину. Функция печени также возвращается к норме.Компенсаторная (викарная) гипертрофия заключается в изменениях в одном из органов при нарушении в другом, относящемся к той же системе органов. Пример — гипертрофия в одной из почек при удалении другой или увеличение лимфатических узлов при удалении селезенки. Изменения способности к такого типа регенерации в зависимости от возраста показаны на рис. 8.80.Последние два способа отличаются местом регенерации, но механизмы их одинаковы: гиперплазия и гипертрофия (рис. 8.81)1.1 Гипертрофия (греч. hyper- + trophe — пища, питание) — увеличение объема и массы органа тела или отдельной его части. Гиперплазия (греч. hyper- + plasis — образование, формирование) — увеличение числа структурных элементов тканей путем их избыточного новообразования. Это не только размножение клеток, но и увеличение цитоплазма-тических ультраструктур (изменяются в первую очередь митохондрии, миофиламенты, эндоплазматический ретикулум, рибосомы). Рис. 8.81. Схема, иллюстрирующая механизмы гипертрофии и гиперплазии: а — норма; б — гиперплазия; в — гипертрофия; г — комбинированное изменениеЭпителизация при заживлении ран с нарушенным эпителиальным покровом идет примерно одинаково, независимо от того, будет далее происходить регенерация органа путем эпиморфоза или нет. Эпидер-мальное заживление раны у млекопитающих в том случае, когда раневая поверхность высыхает с образованием корки, проходит следующим образом (рис. 8.82). Эпителий на краю раны утолщается вследствие увеличения объема клеток и расширения межклеточных пространств. Сгусток фибрина играет роль субстрата для миграции эпидермиса в глубь раны. В мигрирующих эпителиальных клетках нет митозов, одна- Рис. 8.82. Схема некоторых событий, происходящих при эпителизации кожной раны у млекопитающих: а — начало врастания эпидермиса под некротическую ткань, б — срастание эпидермиса и отделение струпа; 1 — соединительная ткань; 2 — эпидермис; 3 — струп; 4 — некротическая тканько они обладают фагоцитарной активностью. Клетки с противоположных краев вступают в контакт. Затем наступает кератинизация раневого эпидермиса и отделение корки, покрывающей рану. К моменту встречи эпидермиса противоположных краев в клетках, расположенных непосредственно вокруг края раны, наблюдается вспышка митозов, которая затем постепенно угасает.Восстановление отдельных мезодермальных тканей, таких как мышечная и скелетная, называют тканевой регенерацией. Для регенерации мышцы важно сохранение хотя бы небольших ее культей на обоих концах, а для регенерации кости необходима надкостница.Таким образом, существует множество различных способов или типов морфогенетических явлений при восстановлении утраченных и поврежденных частей организма. Различия между ними не всегда очевидны, и требуется более глубокое понимание этих процессов.При регенерации не всегда образуется точная копия удаленной структуры. В случае типичной регенерации восстанавливается утраченная часть правильной структуры (гомоморфоз), чего не происходит при атипичной регенерации. Примером последней является появление иной структуры на месте утраченной — гетероморфоз. Она может проявляться в виде гомеозисной регенерации, заключающейся в появлении антенны или конечности на месте глаза у членистоногих. Еще один вариант — гипоморфоз, регенерация с частичным замещением ампутированной структуры. Например, у ящерицы возникает шиловидная структура вместо конечности (рис. 8.83).К атипичной регенерации могут быть отнесены случаи изменения полярности структуры. Так, из короткого фрагмента планарии можно стабильно получать биполярную планарию. Встречается образование дополнительных структур, или избыточная регенерация. После надреза культи при ампутации головного отдела планарии возникает регенерация двух голов или более (рис. 8.84).Изучение регенерации касается не только внешних проявлений. Существует целый ряд аспектов, носящих проблемный и теоретический характер. К ним относятся вопросы регуляции и условий, в которых протекают восстановительные процессы, вопросы происхождения клеток, участвующих в регенерации, способности к регенерации у различных групп животных и особенностей восстановительных процессов у млекопитающих.Установлено, что при регенерации происходят такие процессы, как детерминация, дифференцировка и дифференциация, рост, морфоге- Рис. 8.83. Примеры атипичной регенерации: а — нормальная голова рака; б — формирование антенны вместо глаза; в — образование шиловидной структуры вместо конечности у саламандры. 1 — глаз; 2 — антенна; 3 — место ампутации; 4 — нервный ганглий Рис. 8.84. Примеры атипичной регенерации: а — биполярная планария; б — многоголовая планария, полученная после ампутации головы и нанесения насечек на культюнез, сходные с процессами, имеющими место в эмбриональном развитии. Данные, полученные к настоящему времени, указывают на то, что восстановление утраченных структур, по сути дела, осуществляется на основе той же самой программы развития, которая руководит формированием их у эмбриона, и на основе клеточных и системных механизмов развития. Однако при регенерации все процессы развития идут уже вторично, т.е. в сформированном организме, поэтому восстановление структур имеет ряд отличий и специфичных черт. Несомненно, что в ходе регенерации большое значение принадлежит системным механизмам — межклеточным и межзачатковым взаимодействиям, нервной и гуморальной регуляции. Так, при эпиморфозе конечности тритона сформированный в ходе эпителизации эпидермис стимулирует лизис подлежащих мезодермальных тканей. В его отсутствие или при образовании шрама регенерации не происходит. Клетки под сформированным эпидермисом дедифференцируются и формируют бластему. На этом этапе наблюдаются реципрокные индуктивные влияния между эпидермисом, который формирует апикальную эктодермальную шапочку, и мезодер-мальной бластемой. В ходе эмбрионального развития при формировании конечности осуществлялись сходные взаимодействия между мезодермаль-ной почкой конечности и апикальным эктодермальным гребнем. В ходе дедифференцировки в клетках подавляется активность типо-специфических генов, определяющих специализацию клетки, например генов MRF и Mif5 в мышечных волокнах. Затем активируются гены, необходимые для пролиферации клеток. Один из них msx1. На этой стадии врастающие в бластему нервные отростки и эпидермис продуцируют трофические и ростовые факторы, необходимые для пролиферации и выживания клеток бластемы. Среди них фактор роста фибробластов FGF-10. Этот же фактор необходим для пролиферации самого эпидермиса. Бластема, в свою очередь, синтезирует в ответ нейротрофические факторы, стимулирующие врастание нервов. Нервы нужны для формирования апикальной эктодермальной шапочки. Помимо этого бластема, так же как и апикальная эпидермальная шапочка, продуцирует FGF-8, который стимулирует врастание капилляров. Следует отметить наблюдаемые на этой стадии различия между регенерацией и эмбриональным развитием. Для реализации регенерации необходима иннервация. Без нее может проходить дедифференцировка клеток, но последующее развитие отсутствует. В период эмбрионального морфогенеза конечности (в ходе клеточных дифференцировок) нервы еще не сформированы. Помимо иннервации на ранней стадии регенерации требуется действие ферментов металлопротеиназ. Они разрушают компоненты ма-трикса, что позволяет клеткам разделиться (диссоциировать) и активно пролиферировать. Контактирующие между собой клетки не могут продолжать регенерацию и отвечать на действие ростовых факторов. Таким образом, в ходе регенерации наблюдаются все варианты межклеточных взаимодействий: путем выделения паракринных факторов, диффундирующих от одной клетки к другой, взаимодействия через матрикс и при непосредственном контакте клеточных поверхностей. В стадии дедифференцировки в клетках культи экспрессируются гомеозисные гены HoxD8 и HoxDlO, а с началом дифференцировки — гены HoxD9 и HoxD13. Как было показано в п. 8.3.4, эти же гены активно транскрибируются и в эмбриональном морфогенезе конечности. Важно отметить, что в ходе регенерации утрачивается дифферен-цировка клеток, а их детерминация сохраняется. Уже на стадии недифференцированной бластемы закладываются основные черты регенерирующей конечности. При этом не требуется активация генов, обеспечивающих спецификацию конечности (Tbx-5 для передней и Tbx-4 для задней). Конечность формируется в зависимости от локализации бластемы. Ее развитие происходит так же, как и в эмбриогенезе: сначала проксимальные отделы, а затем дистальные. Проксимально-дистальный градиент, от которого зависит, какие части растущего зачатка станут плечом, какие — предплечьем, а какие — кистью, задается градиентом белка Prod 1. Он локализован на поверхности клеток бластемы и его концентрация выше у основания конечности. Этот белок играет роль рецептора, а сигнальной молекулой (лигандом) для него является белок nAG. Он синтезируется шванновскими клетками, окружающими регенерирующий нерв. При отсутствии этого белка, который через лиганд-рецепторное взаимодействие запускает активацию необходимого для развития каскада генов, регенерации не происходит. Это объясняет феномен отсутствия восстановления конечности при перерезке нерва, а также и при врастании в бластему недостаточного количества нервных волокон. Интересно, что если нерв конечности тритона отвести под кожу основания конечности, то образуется дополнительная конечность. Если его отвести к основанию хвоста — стимулируется образование дополнительного хвоста. Отведение нерва на боковую область никаких дополнительных структур не вызывает. Все это привело к созданию концепции регене-рационных полей. Рис. 8.85. Эксперимент с поворотом бластемы конечности (пояснения в тексте)Аналогично процессу эмбриогенеза формируется и передне-задняя ось в поле развивающейся конечности. В формирующемся зачатке появляется зона поляризующей активности, определяющая асимметрию конечности. Повернув конец культи конечности на 180°, можно получить конечность с зеркальным удвоением пальцев (рис. 8.85).Таким образом, справедливо утверждение, что формирование конечности происходит в поле органа, а бластема является саморегулирующейся системой. Наряду с вышесказанным, доказательством этому служат результаты, полученные в серии экспериментов по пересадке бластемы передней конечности на бластему середины бедра (рис. 8.86). При пересадке в регене-рационное поле другой конечности трансплантат располагается в соответствии с полученной позиционной информацией (градиенты веществ): бластема плеча смещается к середине бедра, предплечья — к голени, запястья — к лапке. Развитие трансплантированной бластемы в соответствующую часть передней конечности происходит в соответствии с ее детерминацией, которая определяется уровнем ампутации.Помимо межклеточных и индукционных взаимодействий, которые оказываются менее разнообразными, чем в ходе эмбрионального морфогенеза, на регенерациюзначительное влияние оказывает нервная и гуморальная регуляция. Это вполне объяснимо тем, что регенерация осуществляется в уже сформированном организме, где основными регулирующими механизмами являются именно последние. Среди гуморальных влияний следует остановиться на действии гормонов. Альдостерон, гормоны щитовидной железы и гипофиза оказывают стимулирующее влияние на восстановление утраченных Рис. 8.86. Опыты по пересадке бластемы передней конечности в поле задней (пояснения в тексте)структур. Сходное действие имеют и метаболиты, выделяемые поврежденной тканью и транспортируемые плазмой крови или передающиеся через межклеточную жидкость. Именно поэтому дополнительное повреждение в некоторых случаях ускоряет процесс регенерации. Помимо перечисленного на регенерацию оказывают влияние и другие факторы, среди которых температура, при которой происходит восстановление, возраст животного, функционирование органа, стимулирующее регенерацию, и в определенных ситуациях изменение электрического заряда в регенерате. Установлено, что в конечности амфибий после ампутации и в процессе регенерации происходят реальные изменения электрической ак- тивности. При проведении электрического тока через ампутированную конечность у взрослых шпорцевых лягушек наблюдается усиление регенерации передних конечностей. В регенератах увеличивается количество нервной ткани, из чего делается вывод, что электрический ток стимулирует врастание нервов в края конечностей, в норме не регенерирующих. Попытки стимулировать подобным образом восстановление конечностей у млекопитающих оказались безуспешными. Под действием электрического тока или при сочетании действия электрического тока с фактором роста нервов удавалось получить у крысы только разрастание скелетной ткани в виде хрящевых и костных мозолей, которые не походили на нормальные элементы скелета конечностей. Один из наиболее интригующих в теории регенерации — вопрос об ее клеточных источниках. Откуда берутся или как возникают недифференцированные клетки бластемы, морфологически сходные с мезен-химными? В настоящее время говорят о трех возможных источниках регенерации. Первый — это дедифференцированные клетки, второй — региональные стволовые клетки и третий — стволовые клетки из других структур, мигрировавшие к месту регенерации. Большинство исследователей признают дедифференцировку и метаплазию при регенерации хрусталика у амфибий. Теоретическое значение этой проблемы заключается в допущении возможности или невозможности изменений клеткой ее программы до такой степени, что она возвращается в состояние, когда снова способна делиться и репро-граммировать свой синтетический аппарат. Наличие региональных стволовых клеток установлено к настоящему времени во многих тканях: в мышцах, кости, эпидермисе кожи, печени, сетчатке и других. Такие клетки обнаружены даже в нервной ткани — в определенных зонах головного мозга. Во многих случаях считают, что источником, из которого образуются дифференцированные клетки в ходе регенерации, являются именно они (регенеративная медицина, регенеративная ветеринария). Предполагается, что по мере увеличения возраста особи численность популяций региональных стволовых клеток сокращается. Если же в органе не хватает своих региональных стволовых клеток, то в него могут мигрировать клетки из других и дать начало нужной ткани. Недавно показано, что стволовые клетки, изолированные из одной взрослой ткани, могут дать начало зрелым клеткам других клеточных линий, независимо от назначения классического зародышевого слоя. Так, эндотелий крупных магистральных артерий не имеет собственных запасов стволовых клеток. Его обновление происходит за счет стволовых клеток костного мозга, поступающих в кровоток. Однако сравнительная неэффективность подобных преобразований in vivo (в организме), даже при наличии повреждения ткани, ставит вопрос о том, имеет ли этот механизм физиологическое значение.Интересно, что среди взрослых стволовых клеток способность к перемене линий наиболее велика у стволовых клеток, которые могут быть культивируемы в среде в течение длительного времени.Если удастся решить вопрос трансформации клеточных линий, то вполне возможным станет использование этих технологий в репаратив-ной медицине для лечения широкого круга болезней. Однако, несмотря на достижения биологии последних лет, в проблеме регенерации еще остается очень много нерешенных вопросов.
Регенерация (в патологии) - это восстановление целости тканей, нарушенной каким-либо болезненным процессом или внешним травмирующим воздействием. Восстановление происходит за счет соседних клеток, заполнения молодыми клетками дефекта и последующего превращения их в зрелую ткань. Такая форма носит название репаративной (возмещающей) регенерации. При этом возможны два варианта регенерации: 1) убыль возмещается тканью того же вида, что и погибшая (полная регенерация); 2) убыль замещается молодой соединительной (грануляционной) тканью, превращающейся в рубцовую (неполная регенерация), что является не регенерацией в собственном смысле, а заживлением тканевого дефекта.
Регенерация предшествует освобождение данного участка от погибших клеток путем ферментативного их расплавления и всасывания в лимфу или кровь или путем (см.). Продукты расплавления являются одним из стимуляторов размножения соседних клеток. Во многих органах и системах существуют участки, клетки которых являются источником размножения клеток при регенерации. Например, в костной системе таким источником является надкостница, клетки которой, размножаясь, образуют вначале остеоидную ткань, в дальнейшем превращающуюся в кость; в слизистых оболочках - клетки глубоколежащих желез (крипты). Регенерация клеток крови происходит в костном мозге и вне его в системе и ее производных (лимфатических узлах, селезенке).
Способностью к регенерации обладают далеко не все ткани и не в одинаковой степени. Так, мышечные клетки сердца не способны к размножению, завершающемуся образованием зрелых мышечных волокон, поэтому всякий дефект мышц миокарда замещается рубцом (в частности, после инфаркта). При гибели ткани мозга (после кровоизлияния, артериосклеротического размягчения) дефект не замещается нервной тканью, а образуется киота.
Иногда возникающая при регенерации ткань по строению отличается от исходной (атипическая регенерация) или объем ее превышает объем погибшей ткани (гиперрегенерация). Такое течение регенерационного процесса может привести к возникновению опухолевого роста.
Регенерация (лат. regenerate - возрождение, восстановление) - восстановление анатомической целости органа или ткани после гибели структурных элементов.
В физиологических условиях процессы регенерации происходят непрерывно с различной интенсивностью в разных органах и тканях соответственно интенсивности отживания клеточных элементов данного органа или ткани и замещения их новообразованными. Непрерывно замещаются форменные элементы крови, клетки покровного эпителия кожи, слизистых оболочек желудочно-кишечного тракта, дыхательных путей. Циклические процессы в женской половой сфере приводят к ритмическому отторжению и обновлению эндометрия путем его регенерации.
Все эти процессы являются физиологическим прототипом патологической регенерации (ее называют еще репаративной). Особенности развития, течения и исхода репаративной регенерации определяются размерами гибели ткани и характером патогенных воздействий. Последнее обстоятельство особенно надо иметь в виду, так как условия и причины гибели ткани имеют существенное значение для регенерационного процесса и его исходов. Так, например, особый характер имеют рубцы после ожогов кожи, отличающиеся от рубцов другого происхождения; сифилитические рубцы грубы, приводят к глубоким втяжениям и обезображиванию органа и т. д. В отличие от физиологической регенерации, репаративная регенерация охватывает широкий круг процессов, ведущих к возмещению дефекта, вызванного утратой ткани вследствие ее повреждения. Различают полную репаративную регенерацию - реституцию (замещение дефекта тканью того же вида и той же структуры, что и погибшая) и неполную репаративную регенерацию (заполнение дефекта тканью, обладающей большими пластическими свойствами, чем погибшая, т. е. обычной грануляционной тканью и соединительной тканью с дальнейшим превращением ее в рубцовую). Таким образом, в патологии под регенерацией часто подразумевают заживление.
С понятием регенерации связано также понятие об организации, поскольку в основе обоих процессов лежат общие закономерности новообразования тканей и понятие субституции, т. е. вытеснения и замещения новообразованной тканью ткани предсуществовавшей (например, субституция тромба фиброзной тканью).
Степень полноты регенерации определяется двумя основными факторами: 1) регенерационная потенция данной ткани; 2) объем дефекта и одно- или разнородность видовой принадлежности погибших тканей.
Первый фактор нередко связывают со степенью дифференцировки данной ткани. Однако само понятие дифференцировки и содержание этого понятия являются весьма относительными, и сравнение тканей по этому признаку с установлением количественной градации дифференцированности в функциональном и морфологическом отношениях невозможно. Наряду с тканями, обладающими высокой регенерационной потенцией (например, ткань печени, слизистые оболочки желудочно-кишечного тракта, органы кроветворения и др.), существуют органы с ничтожной потенцией к регенерации, в которых регенерация никогда не завершается полным восстановлением утраченной ткани (например, миокард, ЦНС). Чрезвычайно высокой пластичностью обладают соединительная ткань, элементы стенки мельчайших кровеносных и лимфатических сосудов, периферические нервы, ретикулярная ткань и ее производные. Поэтому пластическое раздражение, каковым является травма в широком смысле этого слова (т. е. все формы ее), прежде всего и полнее всего стимулирует рост этих тканей.
Объем погибшей ткани имеет существенное значение для полноты регенерации, и эмпирически более или менее известны количественные границы потери ткани для каждого органа, определяющие степень восстановления. Полагают, что для полноты регенерации значение имеет не только объем как чисто количественная категория, но и комплексное разнообразие погибших тканей (это особенно относится к гибели тканей, вызванной токсикоинфекционными воздействиями). Для объяснения этого факта следует, по-видимому, обратиться к общим закономерностям стимуляции пластических процессов в условиях патологии: стимуляторами являются сами продукты тканевой гибели (гипотетические «некрогормоны», «митогенетические лучи», «трефоны» и т. д.). Одни из них являются специфическими стимуляторами для клеток определенного вида, другие - неспецифическими, стимулирующими наиболее пластические ткани. К неспецифическим стимуляторам относят продукты распада и жизнедеятельности лейкоцитов. Их наличие при реактивном воспалении, развивающемся всегда при гибели не только паренхиматозных элементов, но и сосудоносящей стромы, способствует размножению наиболее пластических элементов - соединительной ткани, т. е. развитию в конечном счете рубца.
Существует общая схема последовательности процессов регенерации независимо от области, где она происходит. В условиях патологии процессы регенерации в узком смысле этого слова и процессы заживления имеют различный характер. Это различие определяется характером гибели ткани и избирательной направленностью действия патогенного фактора. Чистые формы регенерации, т. е. восстановление ткани, идентичной утраченной, наблюдаются в тех случаях, когда под влиянием патогенного воздействия погибают только специфические паренхиматозные элементы органа при условии их высокой регенерирующей потенции. Примером этого является регенерация эпителия канальцев почки, избирательно поврежденного токсическим воздействием; регенерация эпителия слизистых оболочек при десквамации его; регенерация альвеолоцитов легкого при десквамативном катаре; регенерация эпителия кожи; регенерация эндотелия кровеносных сосудов и эндокарда и др. В этих случаях источником регенерации являются оставшиеся клеточные элементы, размножение, созревание и дифференциация которых приводит к полному замещению утраченных паренхиматозных элементов. При гибели сложных структурных комплексов восстановление утраченной ткани идет из особых участков органа, являющихся своеобразными центрами регенерации. В слизистой оболочке кишечника, в эндометрии такими центрами являются железистые крипты. Их размножающиеся клетки покрывают дефект сначала одним слоем недифференцированных клеток, из которых затем дифференцируются железы и восстанавливается структура слизистой. В костной системе таким центром регенерации является надкостница, в покровном плоском эпителии - мальпигиев слой, в системе крови - костный мозг и экстрамедуллярные производные ретикулярной ткани.
Общим законом регенерации является закон развития, согласно которому в процессе новообразования возникают юные недифференцированные клеточные производные, в дальнейшем проходящие этапы морфологической и функциональной дифференцировки вплоть до формирования зрелой ткани.
Гибель участков органа, состоящих из комплекса различных тканей, вызывает реактивное воспаление (см.) по периферии. Это приспособительный акт, так как воспалительная реакция сопровождается гиперемией и повышением тканевого метаболизма, что способствует росту новообразующихся клеток. Помимо того, клеточные элементы воспаления из группы гистофагоцитов являются пластическим материалом для новообразования соединительной ткани.
В патологии нередко анатомическое заживление достигается при помощи грануляционной ткани (см.) - этапа новообразования фиброзного рубца. Грануляционная ткань развивается почти при всякой репаративной регенерации, но степень ее развития и конечные исходы варьируют в очень широких пределах. Иногда это с трудом различимые при микроскопическом исследовании нежные участки фиброзной ткани, иногда грубые плотные тяжи гиалинизированной брадитрофной рубцовой ткани, нередко подвергающейся кальцинозу (см.) и оссификации.
Помимо регенерационной потенции данной ткани, характера ее поражения, объема его, важное значение в регенерационном процессе имеют общие факторы. К ним относят возраст субъекта, характер и особенности питания, общую реактивность организма. При нарушениях иннервации, авитаминозах обычное течение репаративной регенерации извращается, что чаще всего выражается в замедлении процесса регенерации, вялости клеточных реакций. Существует также понятие о фибропластическом диатезе как о конституциональной особенности организма реагировать на различные патогенные раздражения повышенным образованием фиброзной ткани, что проявляется формированием келоида (см.), спаечной болезнью. В клинической практике важно учитывать общие факторы для создания оптимальных условий полноты регенерационного процесса и заживления.
Регенерация является одним из важнейших приспособительных процессов, обеспечивающих восстановление здоровья и продолжения жизни при чрезвычайных обстоятельствах, создаваемых болезнью. Однако, как и любой приспособительный процесс, регенерация на известном этапе и при некоторых путях развития может терять приспособительное значение и сама создавать новые формы патологии. Обезображивающие рубцы, деформирующие орган, резко нарушающие его функцию (например, рубцовое превращение клапанов сердца в исходе эндокардита), создают нередко тяжелую хроническую патологию, требующую специальных лечебных мероприятий. Иногда новообразованная ткань количественно превосходит объем погибшей (суперрегенерация). Помимо того, во всяком регенерате имеются элементы атипизма, резкая выраженность которых является этапом развития опухоли (см.). Регенерация отдельных органов и тканей - см. в соответствующих статьях об органах и тканях.
Общие сведения
Регенерация (от лат. regeneratio - возрождение) - восстановление (возмещение) структурных элементов ткани взамен погибших. В биологическом смысле регенерация представляет собой приспособительный процесс, выработанный в ходе эволюции и присущий всему живому. В жизнедеятельности организма каждое функциональное отправление требует затрат материального субстрата и его восстановления. Следовательно, при регенерации происходит самовоспроизведение живой материи, причем это самовоспроизведение живого отражает принцип ауторегуляции и автоматизации жизненных отправлений (Давыдовский И.В., 1969).
Регенераторное восстановление структуры может происходить на разных уровнях - молекулярном, субклеточном, клеточном, тканевом и органном, однако всегда речь идет о возмещении структуры, которая способна выполнять специализированную функцию. Регенерация - это восстановление как структуры, так и функции. Значение регенераторного процесса - в материальном обеспечении гомеостаза.
Восстановление структуры и функции может осуществляться с помощью клеточных или внутриклеточных гиперпластических процессов. На этом основании различают клеточную и внутриклеточную формы регенерации (Саркисов Д.С., 1977). Для клеточной формы регенерации характерно размножение клеток митотическим и амитотическим путем, для внутриклеточной формы, которая может быть органоидной и внутриорганоидной, - увеличение числа (гиперплазия) и размеров (гипертрофия) ультраструктур (ядра, ядрышек, митохондрий, рибосом, пластинчатого комплекса и т.д.) и их компонентов (см. рис. 5, 11, 15). Внутриклеточная форма регенерации является универсальной, так как она свойственна всем органам и тканям. Однако структурно-функциональная специализация органов и тканей в фило- и онтогенезе «отобрала» для одних преимущественно клеточнуую форму, для других - преимущественно или исключительно внутриклеточную, для третьих - в равной мере обе формы регенерации (табл. 5). Преобладание той или иной формы регенерации в определенных органах и тканях определяется их функциональным назначением, структурно-функциональной специализацией. Необходимость сохранения целостности покровов тела объясняет, например, преобладание клеточной формы регенерации эпителия как кожи, так и слизистых оболочек. Специализированная функция пирамидной клетки головного
мозга, как и мышечной клетки сердца, исключает возможность деления этих клеток и позволяет понять необходимость отбора в фило- и онтогенезе внутриклеточной регенерации как единственной формы восстановления данного субстрата.
Таблица 5. Формы регенерации в органах и тканях млекопитающих (по Саркисову Д.С., 1988)
Эти данные опровергают существовавшие до недавнего времени представления об утрате некоторыми органами и тканями млекопитающих способности к регенерации, о «плохо» и «хорошо» регенерирующих тканях человека, о том, что существует «закон обратной зависимости» между степенью дифференцировки тканей и способностью их к регенерации. В настоящее время установлено, что в ходе эволюции способность к регенерации в некоторых тканях и органах не исчезла, а приняла формы (клеточную или внутриклеточную), соответствующие их структурному и функциональному своеобразию (Саркисов Д.С., 1977). Таким образом, все ткани и органы обладают способностью к регенерации, различны лишь ее формы в зависимости от структурно-функциональной специализации ткани или органа.
Морфогенез регенераторного процесса складывается из двух фаз - пролиферации и дифференцировки. Особенно хорошо эти фазы выражены при клеточной форме регенерации. В фазу пролиферации размножаются молодые, недифференцированные клетки. Эти клетки называют камбиальными (от лат. cambium - обмен, смена), стволовыми клетками и клетками-предшественниками.
Для каждой ткани характерны свои камбиальные клетки, которые отличаются степенью пролиферативной активности и специализации, однако одна стволовая клетка может быть родоначальником нескольких видов
клеток (например, стволовая клетка кроветворной системы, лимфоидной ткани, некоторые клеточные представители соединительной ткани).
В фазу дифференцировки молодые клетки созревают, происходит их структурно-функциональная специализация. Та же смена гиперплазии ультраструктур их дифференцировкой (созреванием) лежит в основе механизма внутриклеточной регенерации.
Регуляция регенераторного процесса. Среди регуляторных механизмов регенерации различают гуморальные, иммунологические, нервные, функциональные.
Гуморальные механизмы реализуются как в клетках поврежденных органов и тканей (внутритканевые и внутриклеточные регуляторы), так и за их пределами (гормоны, поэтины, медиаторы, факторы роста и др.). К гуморальным регуляторам относят кейлоны (от греч. chalaino - ослаблять) - вещества, способные подавлять деление клеток и синтез ДНК; они обладают тканевой специфичностью. Иммунологические механизмы регуляции связаны с «регенерационной информацией», переносимой лимфоцитами. В связи с этим следует заметить, что механизмы иммунологического гомеостаза определяют и структурный гомеостаз. Нервные механизмы регенераторных процессов связаны прежде всего с трофической функцией нервной системы, а функциональные механизмы - с функциональным «запросом» органа, ткани, который рассматривается как стимул к регенерации.
Развитие регенераторного процесса во многом зависит от ряда общих и местных условий, или факторов. К общим следует отнести возраст, конституцию, характер питания, состояние обмена и кроветворения, к местным - состояние иннервации, крово- и лимфообращения ткани, пролиферативную активность ее клеток, характер патологического процесса.
Классификация. Различают три вида регенерации: физиологическую, репаративную и патологическую.
Физиологическая регенерация совершается в течение всей жизни и характеризуется постоянным обновлением клеток, волокнистых структур, основного вещества соединительной ткани. Нет таких структур, которые не подвергались бы физиологической регенерации. Там, где доминирует клеточная форма регенерации, имеет место обновление клеток. Так происходит постоянная смена покровного эпителия кожи и слизистых оболочек, секреторного эпителия экзокринных желез, клеток, выстилающих серозные и синовиальные оболочки, клеточных элементов соединительной ткани, эритроцитов, лейкоцитов и тромбоцитов крови и т.д. В тканях и органах, где клеточная форма регенерации утрачена, например в сердце, головном мозге, происходит обновление внутриклеточных структур. Наряду с обновлением клеток и субклеточных структур постоянно совершается биохимическая регенерация, т.е. обновление молекулярного состава всех компонентов тела.
Репаративная или восстановительная регенерация наблюдается при различных патологических процессах, ведущих к повреждению клеток и тка-
ней. Механизмы репаративной и физиологической регенерации едины, репаративная регенерация - это усиленная физиологическая регенерация. Однако в связи с тем, что репаративная регенерация побуждается патологическими процессами, она имеет качественные морфологические отличия от физиологической. Репаративная регенерация может быть полной и неполной.
Полная регенерация, или реституция, характеризуется возмещением дефекта тканью, которая идентична погибшей. Она развивается преимущественно в тканях, где преобладает клеточная регенерация. Так, в соединительной ткани, костях, коже и слизистых оболочках даже относительно крупные дефекты органа могут путем деления клеток замещаться тканью, идентичной погибшей. При неполной регенерации, или субституции, дефект замещается соединительной тканью, рубцом. Субституция характерна для органов и тканей, в которых преобладает внутриклеточная форма регенерации, либо она сочетается с клеточной регенерацией. Поскольку при регенерации происходит восстановление структуры, способной к выполнению специализированной функции, смысл неполной регенерации не в замещении дефекта рубцом, а в компенсаторной гиперплазии элементов оставшейся специализированной ткани, масса которой увеличивается, т.е. происходит гипертрофия ткани.
При неполной регенерации, т.е. заживлении ткани рубцом, возникает гипертрофия как выражение регенераторного процесса, поэтому ее называют регенерационной, в ней - биологический смысл репаративной регенерации. Регенераторная гипертрофия может осуществляться двумя путями - с помощью гиперплазии клеток или гиперплазии и гипертрофии клеточных ультраструктур, т.е. гипертрофии клеток.
Восстановление исходной массы органа и его функции за счет преимущественно гиперплазии клеток происходит при регенерационной гипертрофии печени, почек, поджелудочной железы, надпочечников, легких, селезенки и др. Регенерационная гипертрофия за счет гиперплазии клеточных ультраструктур характерна для миокарда, головного мозга, т.е. тех органов, где преобладает внутриклеточная форма регенерации. В миокарде, например, по периферии рубца, заместившего инфаркт, размеры мышечных волокон значительно увеличиваются, т.е. они гипертрофируются в связи с гиперплазией их субклеточных элементов (рис. 81). Оба пути регенерационной гипертрофии не исключают друг друга, а, наоборот, нередко сочетаются. Так, при регенерационной гипертрофии печени происходит не только увеличение числа клеток в сохранившейся после повреждения части органа, но и гипертрофия их, обусловленная гиперплазией ультраструктур. Нельзя исключить того, что в мышце сердца регенерационная гипертрофия может протекать не только в виде гипертрофии волокон, но и путем увеличения числа составляющих их мышечных клеток.
Восстановительный период обычно не ограничивается только тем, что в поврежденном органе развертывается репаративная регенерация. Если
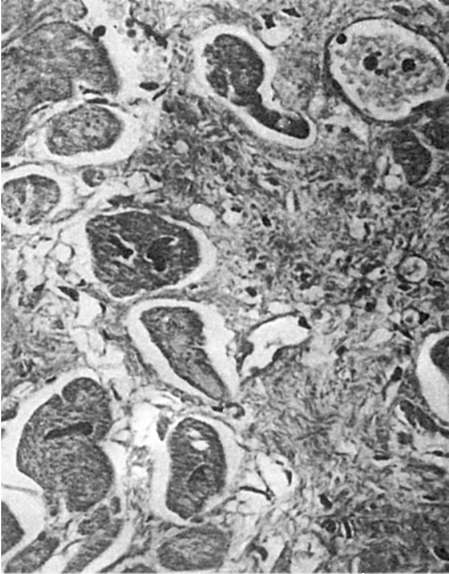 Рис. 81.
Регенерационная гипертрофия миокарда. По периферии рубца расположены гипертрофированные мышечные волокна
Рис. 81.
Регенерационная гипертрофия миокарда. По периферии рубца расположены гипертрофированные мышечные волокна
воздействие патогенного фактора прекращается до гибели клетки, происходит постепенное восстановление поврежденных органелл. Следовательно, проявления репаративной реакции должны быть расширены за счет включения восстановительных внутриклеточных процессов в дистрофически измененных органах. Общепринятое мнение о регенерации только как о завершающем этапе патологического процесса малооправданно. Репаративная регенерация не местная, а общая реакция организма, охватывающая различные органы, но реализующаяся в полной мере лишь в том или ином из них.
О патологической регенерации говорят в тех случаях, когда в результате тех или иных причин имеется извращение регенераторного процесса, нарушение смены фаз пролиферации
и дифференцировки. Патологическая регенерация проявляется в избыточном или недостаточном образовании регенерирующей ткани (гипер- или гипорегенерация), а также в превращении в ходе регенерации одного вида ткани в другой [метаплазия - см. Процессы приспособления (адаптации) и компенсации]. Примерами могут служить гиперпродукция соединительной ткани с образованием келоида, избыточная регенерация периферических нервов и избыточное образование костной мозоли при срастании перелома, вялое заживление ран и метаплазия эпителия в очаге хронического воспаления. Патологическая регенерация обычно развивается при нарушениях общих и местных условий регенерации (нарушение иннервации, белковое и витаминное голодание, хроническое воспаление и т.д.).
Регенерация отдельных тканей и органов
Репаративная регенерация крови отличается от физиологической прежде всего своей большей интенсивностью. При этом активный красный костный мозг появляется в длинных трубчатых костях на месте жирового костного мозга (миелоидное превращение жирового костного мозга). Жировые клетки вытесняются растущими островками кроветворной ткани, которая заполняет костномозговой канал и выглядит сочной, темнокрасной. Кроме того, кроветворение начинает происходить вне костного мозга - внекостномозговое, или экстрамедуллярное, кроветворение. Оча-
ги экстрамедуллярного (гетеротопического) кроветворения в результате выселения из костного мозга стволовых клеток появляются во многих органах и тканях - селезенке, печени, лимфатических узлах, слизистых оболочках, жировой клетчатке и т.д.
Регенерация крови может быть резко угнетена (например, при лучевой болезни, апластической анемии, алейкии, агранулоцитозе) или извращена (например, при злокачественной анемии, полицитемии, лейкозе). В кровь при этом поступают незрелые, функционально неполноценные и быстро разрушающиеся форменные элементы. В таких случаях говорят о патологической регенерации крови.
Репаративные возможности органов кроветворной и иммунокомпетентной системы неоднозначны. Костный мозг обладает очень высокими пластическими свойствами и может восстанавливаться даже при значительных повреждениях. Лимфатические узлы хорошо регенерируют только в тех случаях, когда сохраняются связи приносящих и выносящих лимфатических сосудов с окружающей их соединительной тканью. Регенерация ткани селезенки при повреждении бывает, как правило, неполной, погибшая ткань замещается рубцом.
Регенерация кровеносных и лимфатических сосудов протекает неоднозначно в зависимости от их калибра.
Микрососуды обладают большей способностью регенерировать, чем крупные сосуды. Новообразование микрососудов может происходить путем почкования или аутогенно. При регенерации сосудов путем почкования (рис. 82) в их стенке появляются боковые выпячивания за счет усиленно делящихся эндотелиальных клеток (ангиобласты). Образуются тяжи из эндотелия, в которых возникают просветы и в них поступает кровь или лимфа из «материнского» сосуда. Другие элементы: сосудистой стенки образуются за счет дифференцировки эндотелия и окружающих сосуд соединительнотканных клеток, В сосудистую стенку врастают нервные волокна из предсуществующих нервов. Аутогенное новообразование сосудов состоит в том, что в соединительной ткани появляются очаги недифференцированных клеток. В этих очагах возникают щели, в которые открываются предсуществующие капилляры и изливается кровь. Молодые клетки соединительной ткани, дифференцируясь, образуют эндотелиальную выстилку и другие элементы стенки сосуда.
 Рис. 82.
Регенерация сосудов путем почкования
Рис. 82.
Регенерация сосудов путем почкования
Крупные сосуды не обладают достаточными пластическими свойствами. Поэтому при повреждении их стенки восстанавливаются лишь структуры внутренней оболочки, ее эндотелиальная выстилка; элементы средней и наружной оболочек обычно замещаются соединительной тканью, что ведет нередко к сужению или облитерации просвета сосуда.
Регенерация соединительной ткани начинается с пролиферации молодых мезенхимальных элементов и новообразования микрососудов. Образуется молодая, богатая клетками и тонкостенными сосудами соединительная ткань, которая имеет характерный вид. Это - сочная темнокрасная ткань с зернистой, как бы усыпанной крупными гранулами поверхностью, что явилось основанием назвать ее грануляционной тканью. Гранулы представляют собой выступающие над поверхностью петли новообразованных тонкостенных сосудов, которые составляют основу грануляционной ткани. Между сосудами много недифференцированных лимфоцитоподобных клеток соединительной ткани, лейкоцитов, плазматических клеток и лаброцитов (рис. 83). В дальнейшем происходит созревание грануляционной ткани, в основе которой лежит дифференцировка клеточных элементов, волокнистых структур, а также сосудов. Число гематогенных элементов уменьшается, а фибробластов - увеличивается. В связи с синтезом фибробластами коллагена в межклеточных пространствах образуются аргирофильные (см. рис. 83), а затем и коллагеновые волокна. Синтез фибробластами гликозаминогликанов служит образованию
основного вещества соединительной ткани. По мере созревания фибробластов количество коллагеновых волокон увеличивается, они группируются в пучки; одновременно уменьшается количество сосудов, они дифференцируются в артерии и вены. Созревание грануляционной ткани завершается образованием грубоволокнистой рубцовой ткани.
Новообразование соединительной ткани происходит не только при ее повреждении, но и при неполной регенерации других тканей, а также при организации (инкапсуляции), заживлении ран, продуктивном воспалении.
Созревание грануляционной ткани может иметь те или иные отклонения. Воспаление, развивающееся в грануляционной ткани, приводит к задержке ее созревания,
 Рис. 83.
Грануляционная
ткань. Между тонкостенными сосудами много недифференцированных клеток
соединительной ткани и аргирофильных волокон. Импрегнация серебром
Рис. 83.
Грануляционная
ткань. Между тонкостенными сосудами много недифференцированных клеток
соединительной ткани и аргирофильных волокон. Импрегнация серебром
а чрезмерная синтетическая активность фибробластов - к избыточному образованию коллагеновых волокон с последующим резко выраженным их гиалинозом. В таких случаях возникает рубцовая ткань в виде опухолевидного образования синюшно-красного цвета, которая возвышается над поверхностью кожи в виде келоида. Келоидные рубцы образуются после различных травматических поражений кожи, особенно после ожогов.
Регенерация жировой ткани происходит за счет новообразования соединительнотканных клеток, которые превращаются в жировые (адипозоциты) путем накопления в цитоплазме липидов. Жировые клетки складываются в дольки, между которыми располагаются соединительнотканные прослойки с сосудами и нервами. Регенерация жировой ткани может происходить также из ядросодержащих остатков цитоплазмы жировых клеток.
Регенерация костной ткани при переломе костей в значительной мере зависит от степени разрушения кости, правильной репозиции костных отломков, местных условий (состояние кровообращения, воспаление и т.д.). При неосложненном костном переломе, когда костные отломки неподвижны, может происходить первичное костное сращение (рис. 84). Оно начинается с врастания в область дефекта и гематомы между отломками кости молодых мезенхимальных элементов и сосудов. Возникает так называемая предварительная соединительнотканная мозоль, в которой сразу же начинается образование кости. Оно связано с активацией и пролиферацией остеобластов в зоне повреждения, но прежде всего в периостате и эндостате. В остеогенной фиброретикулярной ткани появляются малообызвествленные костные балочки, число которых нарастает.
Образуется предварительная костная мозоль. В дальнейшем она созревает и превращается в зрелую пластинчатую кость - так образуется
 Рис. 84.
Первичное костное сращение. Интермедиарная костная мозоль (показана стрелкой), спаивающая отломки кости (по Г.И. Лаврищевой)
Рис. 84.
Первичное костное сращение. Интермедиарная костная мозоль (показана стрелкой), спаивающая отломки кости (по Г.И. Лаврищевой)
окончательная костная мозоль, которая по своему строению отличается от костной ткани лишь беспорядочным расположением костных перекладин. После того как кость начинает выполнять свою функцию и появляется статическая нагрузка, вновь образованная ткань с помощью остеокластов и остеобластов подвергается перестройке, появляется костный мозг, восстанавливаются васкуляризация и иннервация. При нарушении местных условий регенерации кости (расстройство кровообращения), подвижности отломков, обширных диафизарных переломах происходит вторичное костное сращение (рис. 85). Для этого вида костного сращения характерно образование между костными отломками сначала хрящевой ткани, на основе которой строится костная ткань. Поэтому при вторичном костном сращении говорят о предварительной костно-хрящевой мозоли, которая со временем превращается в зрелую кость. Вторичное костное сращение по сравнению с первичным встречается значительно чаще и занимает больше времени.
При неблагоприятных условиях регенерация костной ткани может быть нарушена. Так, при инфицировании раны регенерация кости задерживается. Костные осколки, которые при нормальном течении регенераторного процесса выполняют функцию каркаса для новообразованной костной ткани, в условиях нагноения раны поддерживают воспаление, что тормозит регенерацию. Иногда первичная костно-хрящевая мозоль не дифференцируется в костную. В этих случаях концы сломанной кости остаются подвижными, образуется ложный сустав. Избыточная продукция костной ткани в ходе регенерации приводит к появлению костных выростов - экзостозов.
Регенерация хрящевой ткани в отличие от костной происходит обычно неполно. Лишь небольшие дефекты ее могут замещаться новообразованной тканью за счет камбиальных элементов надхрящницы - хондробластов. Эти клетки создают основное вещество хряща, затем превращаются в зрелые хрящевые клетки. Крупные дефекты хряща замещаются рубцовой тканью.
Регенерация мышечной ткани, ее возможности и формы различны в зависимости от вида этой ткани. Гладкие мьшщы, клетки которых обладают способностью к митозу и амитозу, при незначительных дефектах могут регенерировать достаточно полно. Значительные участки повреждения гладких мышц замещаются рубцом, при этом сохранившиеся мышечные волокна подвергаются гипертрофии. Новообразование гладких мышечных волокон может происходить путем превращения (метаплазии) элементов соединительной ткани. Так образуются пучки гладких мышечных волокон в плевральных спайках, в подвергающихся организации тромбах, в сосудах при их дифференцировке.
Поперечнополосатые мышцы регенерируют лишь при сохранении сарколеммы. Внутри трубок из сарколеммы осуществляется регенерация ее органелл, в результате чего появляются клетки, называемые миобластами. Они вытягиваются, число ядер в них увеличивается, в саркоплазме
 Рис. 85.
Вторичное костное сращение (по Г.И. Лаврищевой):
Рис. 85.
Вторичное костное сращение (по Г.И. Лаврищевой):
а - костно-хрящевая периостальная мозоль; участок костной ткани среди хрящевой (микроскопическая картина); б - периостальная костно-хрящевая мозоль (гистотопограмма через 2 мес после операции): 1 - костная часть; 2 - хрящевая часть; 3 - отломки кости; в - периостальная мозоль, спаивающая смещенные отломки кости
дифференцируются миофибриллы, и трубки сарколеммы превращаются в поперечнополосатые мышечные волокна. Регенерация скелетных мышц может быть связана и с клетками-сателлитами, которые располагаются под сарколеммой, т.е. внутри мышечного волокна, и являются камбиальными. В случае травмы клетки-сателлиты начинают усиленно делиться, затем подвергаются дифференцировке и обеспечивают восстановление мышечных волокон. Если при повреждении мышцы целость волокон нарушается, то на концах их разрывов возникают колбообразные выбухания, которые содержат большое число ядер и называются мышечными почками. При этом восстановления непрерывности волокон не происходит. Место разрыва заполняется грануляционной тканью, превращающейся в рубец (мышечная мозоль). Регенерация мышцы сердца при ее повреждении, как и при повреждении поперечнополосатой мускулатуры, заканчивается рубцеванием дефекта. Однако в сохранившихся мышечных волокнах происходит интенсивная гиперплазия ультраструктур, что ведет к гипертрофии волокон и восстановлению функции органа (см. рис. 81).
Регенерация эпителия осуществляется в большинстве случаев достаточно полно, так как он обладает высокой регенераторной способностью. Особенно хорошо регенерирует покровный эпителий. Восстановление многослойного плоского ороговевающего эпителия возможно даже при довольно крупных дефектах кожи. При регенерации эпидермиса в краях дефекта происходит усиленное размножение клеток зародышевого (камбиального), росткового (мальпигиева) слоя. Образующиеся эпителиальные клетки сначала покрывают дефект одним слоем. В дальнейшем пласт эпителия становится многослойным, клетки его дифференцируются, и он приобретает все признаки эпидермиса, включающего в себя ростковый, зернистый блестящий (на подошвах и ладонной поверхности кистей) и роговой слои. При нарушении регенерации эпителия кожи образуются незаживающие язвы, нередко с разрастанием в их краях атипичного эпителия, что может послужить основой для развития рака кожи.
Покровный эпителий слизистых оболочек (многослойный плоский неороговевающий, переходный, однослойный призматический и многоядерный мерцательный) регенерирует таким же образом, как и многослойный плоский ороговевающий. Дефект слизистой оболочки восстанавливается за счет пролиферации клеток, выстилающих крипты и выводные протоки желез. Недифференцированные уплощенные клетки эпителия сначала покрывают дефект тонким слоем (рис. 86), затем клетки принимают форму, свойственную клеточным структурам соответствующей эпителиальной выстилки. Параллельно частично или полностью восстанавливаются и железы слизистой оболочки (например, трубчатые железы кишки, железы эндометрия).
Регенерация мезотелия брюшины, плевры и околосердечной сумки осуществляется путем деления сохранившихся клеток. На поверхности дефекта появляются сравнительно крупные кубические клетки, которые затем уплощаются. При небольших дефектах мезотелиальная выстилка восстанавливается быстро и полно.
Важное значение для восстановления покровного эпителия и мезотелия имеет состояние подлежащей соединительной ткани, так как эпителизация любого дефекта возможна лишь после заполнения его грануляционной тканью.
Регенерация специализированного эпителия органов (печени, поджелудочной железы, почек, желез внутренней секреции, легочных альвеол) осуществляется по типу регенерационной гипертрофии: в участках повреждения ткань замещается рубцом, а по периферии его происходят гиперплазия и гипертрофия клеток паренхимы. В печени участок некроза всегда подвергается рубцеванию, однако в остальной части органа происходит интенсивное новообразование клеток, а также гиперплазия внутриклеточных стуктур, что сопровождается их гипертрофией. В результате этого исходная масса и функция органа быстро восстанавливаются. Регенераторные возможности печени почти безграничны. В поджелудочной железе регенераторные процессы хорошо выражены как в экзокринных отделах, так и в панкреатических островках, причем эпителий экзокринных желез становится источником восстановления островков. В почках при некрозе эпителия канальцев происходит размножение сохранившихся нефроцитов и восстановление канальцев, однако лишь при сохранении тубулярной базальной мембраны. При ее разрушении (тубулорексис) эпителий не восстанавливается и каналец замещается соединительной тканью. Не восстанавливается погибший канальцевый эпителий и в том случае, когда одновременно с канальцем погибает сосудистый клубочек. При этом на месте погибшего нефрона разрастается рубцовая соединительная ткань, а окружающие нефроны подвергаются регенерационной гипертрофии. В железах внутренней секреции восстановительные процессы также представлены неполной регенерацией. В легком после удаления отдельных долей в оставшейся части происходит гипертрофия и гиперплазия тканевых элементов. Регенерация специализированного эпителия органов может протекать атипично, что ведет к разрастанию соединительной ткани, структурной перестройке и деформации органов; в таких случаях говорят о циррозе (цирроз печени, нефроцирроз, пневмоцирроз).
Регенерация разных отделов нервной системы происходит неоднозначно. В головном и спинном мозге новообразования ганглиозных клеток не про-
 Рис. 86.
Регенерация эпителия в дне хронической язвы желудка
Рис. 86.
Регенерация эпителия в дне хронической язвы желудка
исходит и при разрушении их восстановление функции возможно лишь за счет внутриклеточной регенерации сохранившихся клеток. Невроглии, особенно микроглии, свойственна клеточная форма регенерации, поэтому дефекты ткани головного и спинного мозга обычно заполняются пролиферирующими клетками невроглии - возникают так называемые глиальные (глиозные) рубцы. При повреждении вегетативных узлов наряду с гиперплазией ультраструктур клеток происходит и их новообразование. При нарушении целости периферического нерва регенерация происходит за счет центрального отрезка, сохранившего связь с клеткой, в то время как периферический отрезок погибает. Размножающиеся клетки шванновской оболочки погибшего периферического отрезка нерва располагаются вдоль него и образуют футляр - так называемый бюнгнеровский тяж, в который врастают регенерирующие осевые цилиндры из проксимального отрезка. Регенерация нервных волокон завершается их миелинизацией и восстановлением нервных окончаний. Регенерационная гиперплазия рецепторов, перицеллюлярных синаптических приборов и эффекторов иногда сопровождается гипертрофией их концевых аппаратов. Если регенерация нерва в силу тех или иных причин нарушается (значительное расхождение частей нерва, развитие воспалительного процесса), то в месте его перерыва образуется рубец, в котором беспорядочно располагаются регенерировавшие осевые цилиндры проксимального отрезка нерва. Аналогичные разрастания возникают на концах перерезанных нервов в культе конечности после ее ампутации. Такие разрастания, образованные нервными волокнами и фиброзной тканью, называются ампутационными невромами.
Заживление ран
Заживление ран протекает по законам репаративной регенерации. Темпы заживления ран, его исходы зависят от степени и глубины раневого повреждения, структурных особенностей органа, общего состояния организма, применяемых методов лечения. По И.В. Давыдовскому, выделяют следующие виды заживления ран: 1) непосредственное закрытие дефекта эпителиального покрова; 2) заживление под струпом; 3) заживление раны первичным натяжением; 4) заживление раны вторичным натяжением, или заживление раны через нагноение.
Непосредственное закрытие дефекта эпителиального покрова - это простейшее заживление, заключающееся в наползании эпителия нд поверхностный дефект и закрытии его эпителиальным слоем. Наблюдаемое на роговице, слизистых оболочках заживление под струпом касается мелких дефектов, на поверхности которых быстро возникает подсыхающая корочка (струп) из свернувшейся крови и лимфы; эпидермис восстанавливается под корочкой, которая отпадает через 3-5 сут после ранения.
Заживление первичным натяжением (per rimamm intentionem) наблюдается в ранах с повреждением не только кожи, но и подлежащей ткани,
причем края раны ровные. Рана заполняется свертками излившейся крови, что предохраняет края раны от дегидратации и инфекции. Под влиянием протеолитических ферментов неитрофилов происходит частичный лизис свертка крови, тканевого детрита. Нейтрофилы погибают, на смену им приходят макрофаги, которые фагоцитируют эритроциты, остатки поврежденной ткани; в краях раны обнаруживается гемосидерин. Часть содержимого раны удаляется в первый день ранения вместе с экссудатом самостоятельно или при обработке раны - первичное очищение. На 2-3-и сутки в краях раны появляются растущие навстречу друг другу фибробласты и новообразованные капилляры, появляется грануляционная ткань, пласт которой при первичном натяжении не достигает больших размеров. К 10-15-м суткам она полностью созревает, раневой дефект эпителизируется и рана заживает нежным рубчиком. В хирургической ране заживление первичным натяжением ускоряется в связи с тем, что ее края стягиваются нитями шелка или кетгута, вокруг которых скапливаются рассасывающие их гигантские клетки инородных тел, не мешающие заживлению.
Заживление вторичным натяжением (per secundam intentionem), или заживление через нагноение (или заживление посредством гранулирования - per granulationem), наблюдается обычно при обширных ранениях, сопровождающихся размозжением и омертвением тканей, проникновении в рану инородных тел, микробов. На месте раны возникают кровоизлияния, травматический отек краев раны, быстро появляются признаки демаркационного гнойного воспаления на границе с омертвевшей тканью, расплавление некротических масс. В течение первых 5-6 сут происходит отторжение некротических масс - вторичное очищение раны, и в краях раны начинает развиваться грануляционная ткань. Грануляционная ткань, выполняющая рану, состоит из 6 переходящих друг в друга слоев (Аничков Н.Н., 1951): поверхностный лейкоцитарно-некротический слой; поверхностный слой сосудистых петель, слой вертикальных сосудов, созревающий слой, слой горизонально расположенных фибробластов, фиброзный слой. Созревание грануляционной ткани при заживлении раны вторичным натяжением сопровождается регенерацией эпителия. Однако при этом виде заживления раны на месте ее всегда образуется рубец.
Регенерация - восстановление организмом утраченных или поврежденных органов и тканей, а также восстановление целого организма из его части. В большей
степени присуща растениям и беспозвоночным животным, в меньшей - позвоночным. Регенерацию можно вызвать
экспериментально.
Регенерация направлена на восстановление поврежденных структурных элементов и регенерационные процессы могут
осуществляться на разных уровнях:
а) молекулярный
б) субклеточный
в) клеточный - размножение клеток митозом и амитотическим путем
г) тканевой
д) органный.
Виды регенерации:
7. Физиологическая - обеспечивает функционирование органов и систем в обычных условиях. Во всех органах происходит физиологическая регенерация, но в каких-то больше, в других - меньше.
2. Репаративная (восстановительная) - возникает в связи с патологическим процессов, который приводит к повреждению ткани (это усиленная физиологическая регенерация)
а) полная регенерация (реституция) - на месте повреждения ткани возникает точно такая же ткань
б) неполная регенерация (субституция) - на месте погибшей ткани возникает соединительная ткань. Например, в сердце при инфаркте миокарда происходит некроз, который замещается соединительной тканью.
Смысл неполной регенерации: вокруг соединительной ткани возникает регенерационная гипертрофия, которая и
обеспечивает сохранение функции поврежденного органа.
Регенерационная гипертрофия осуществляется за счет:
а) гиперплазии клеток (избыточное образование)
б) гипертрофии клеток (увеличение органа в объеме и массе).
Регенерационная гипертрофия в миокарде осуществляется за счет гиперплазии внутриклеточных структур.
Формы регенерации.
1. Клеточная - происходит размножение клеток митотическим и амитотическим путем. Она существует в костной ткани, эпидермисе, слизистой ЖКТ, слизистой дыхательных путей, слизистой мочеполовой системы, эндотелий, мезотелий, рыхлая соединительная ткань, кроветворная система. В этих органах и тканях возникает полная регенерация (точно такая же ткань).
2. Внутриклеточная - происходит гиперплазия внутриклеточных структур. Миокард, скелетные мышцы (преимущественно), ганглиозные клетки ЦНС (исключительно).
3. Клеточные и внутриклеточные формы. Печень, почки, легкие, гладкие мышцы, вегетативная нервная система, поджелудочная железа, эндокринная система. Обычно возникает неполная регенерация.
Регенерация соединительной ткани.
Этапы:
1. Образование грануляционной ткани. Постепенно идет вытеснение сосудов и клеток с образованием волокон. Фибробласты - фиброциты, которые продуцируют волокна.
2. Образование зрелой соединительной ткани. Регенерация крови
1. Физиологическая регенерация. В костном мозге.
2. Репаративная регенерация. Возникает при анемиях, лейкопениях, тромбоцитопениях. Появляются экстрамедуллярные очаги кроветворения (в печени, селезенке, лимфатических узлах, желтый костный мозг участвует в кроветворении).
3. Патологическая регенерация. При лучевой болезни, лейкозах. В органах кроветворения образуются незрелые
кроветворные элементы (властные клетки).
Вопрос 16
ГОМЕОСТАЗ.
Гомеостаз – поддержание постоянства внутренней среды организма в непрерывно изменяющихся условиях внешней среды. Т.к. организм – многоуровневый саморегулирующийся объект, его можно рассматривать с точки зрения кибернетики. Тогда, организм – сложная многоуровневая саморегулирующаяся система с множеством переменных.
Переменные входа:
Причина;
Раздражение.
Переменные выхода:
Реакция;
Следствие.
Причина – отклонение от нормы реакции в организме. Решающая роль принадлежит обратной связи. Существует положительная и отрицательная обратная связь.
Отрицательная обратная связь уменьшает действие входного сигнала на выходной. Положительная обратная связь увеличивает действие входного сигнала на выходной эффект действия.
Живой организм – ультрастабильная система, осуществляющая поиск наиболее оптимального устойчивого состояния, которое обеспечивается адаптациями.
Вопрос 18:
ПРОБЛЕМЫ ТРАНСПЛАНТАЦИИ.
Трансплантация- пересадка тканей и органов.
Трансплантация у животных и человека - приживление органов или участков отдельных тканей для замещения дефектов, стимулирования регенерации, при косметических операциях, а также в целях эксперимента и тканевой терапии.
Аутотрансплантация- пересадка тканей в пределах одного организма Аллотрансплантация- пересадка между организмами одного вида. Ксенотрансплантация- пересадка между различными видами.
Вопрос 19
Хронобиология - раздел биологии, изучающий биологические ритмы, протекание различных биологических процессов
(преимущественно циклических) во времени.
Биологические ритмы - (биоритмы), циклические колебания интенсивности и характера биологических процессов и явлений. Одни биологические ритмы относительно самостоятельны (напр., частота сокращений сердца, дыхания), другие связаны с приспособлением организмов к геофизическим циклам - суточным (напр., колебания интенсивности деления клеток, обмена веществ, двигательной активности животных), приливным (напр., биологические процессы у организмов, связанные с уровнем морских приливов), годичным (изменение численности и активности животных, роста и развития растений и др.). Наука о биологических ритмах - хронобиология.
Вопрос 20
ФИЛОГЕНЕЗ СКЕЛЕТА
Скелет рыб состоит из черепа, позвоночника, скелета непарных, парных плавников и их поясов. В туловищном отделе к поперечным отросткам тела причленяются ребра. Позвонки сочленяются друг с другом при помощи суставных отростков, обеспечивая изгиб преимущественно в горизонтальной плоскости.
Скелет земноводных, как и у всех позвоночных, состоит из черепа, позвоночника, скелета конечностей и их поясов. Череп почти сплошь хрящевой (рис. 11.20). Он подвижно сочленен с позвоночником. Позвоночник содержит девять позвонков, объединенных в три отдела: шейный (1 позвонок), туловищный (7 позвонков), крестцовый (1 позвонок), а все хвостовые позвонки срослись, образовав единую косточку - уростиль. Ребра отсутствуют. Плечевой пояс включает типичные для наземных позвоночных кости: парные лопатки, вороньи кости (коракоиды), ключицы и непарную грудину. Он имеет вид полукольца, лежащего в толще туловищной мускулатуры, т. е. не соединен с позвоночником. Тазовый пояс образован двумя тазовыми костями, образованными тремя парами подвздошных, седалищных и лобковых костей, сросшихся между собой. Длинные подвздошные кости причле-нены к поперечным отросткам крестцового позвонка. Скелет свободных конечностей построен по типу системы многочленных рычагов, подвижно соединенных шаровидными суставами. В составе передней конечности. выделяют плечо, предплечье и кисть.
Тело ящерицы подразделено на голову, туловище и хвост. В туловищном отделе хорошо выражена шея. Все тело покрыто роговыми чешуями, а голова и брюхо - крупными щитками. Конечности ящерицы хорошо развиты и вооружены пятью пальцами с когтями. Плечевые и бедренные кости расположены параллельно поверхности земли, вследствие чего тело провисает и касается земли (отсюда и название класса). Шейный отдел позвоночника состоит из восьми позвонков, первый из них подвижно соединен как с черепом, так и со вторым позвонком, что обеспечивает головному отделу большую свободу движений. Позвонки пояснично-грудного отдела несут ребра, часть которых соединена с грудиной, в результате чего образуется грудная клетка. Крестцовые позвонки обеспечивают более прочное, чем у земноводных, соединение с костями таза.
Скелет млекопитающих по строению в основном сходен со скелетом наземных позвоночных, однако имеются некоторые различия: число шейных позвонков постоянно и равно семи, череп более объемный, что связано с большими размерами головного мозга. Кости черепа срастаются довольно поздно, что обеспечивает возможность увеличения головного мозга по мере роста животного. Конечности млекопитающих построены по пятипалому типу, характерному для наземных позвоночных.
Вопрос 21
ФИЛОГЕНЕЗ КРОВЕНОСНОЙ СИСТЕМЫ
Кровеносная система рыб заткнута. Сердце двухкамерное, состоящее из предсердия и желудочка. Венозная кровь из желудочка сердца поступает в брюшную аорту, несущую ее к жабрам, где она обогащается кислородом и освобождается от углекислого газа. Оттекающая от жабр артериальная кровь собирается в спинную аорту, которая расположена вдоль тела под позвоночником. От спинной аорты к различным органам рыбы отходят многочисленные артерии. В них артерии распадаются на сеть тончайших, капилляров, через стенки которых кровь отдает кислород и обогащается углекислым газом. Венозная кровь собирается в вены и по ним поступает в предсердие, а из него желудочек. Следовательно, у рыб один круг кровообращения.
Кровеносная система земноводных представлена трехкамерным сердцем, состоящим из двух предсердий и желудочка, и двух кругов кровообращения - большого (туловищного) и малого (легочного). Малый круг кровообращения начинается в желудочке, включает сосуды легких и завершается в левом предсердии. Большой круг начинается также в желудочке. Кровь, пройдя по сосудам всего тела, возвращается в правое предсердие. Таким образом, в левое предсердие попадает артериальная кровь из легких, а в правое - венозная кровь со всего тела. В правое предсердие попадает и артериальная кровь, оттекающая от кожи. Так благодаря появлению легочного круга кровообращения в сердце земноводных попадает и артериальная кровь. Несмотря на то что в желудочек поступает артериальная и венозная кровь, полного перемешивания крови не происходит благодаря наличию карманов и неполных перегородок. Благодаря им при выходе из желудочка артериальная кровь по сонным артериям поступает в головной отдел, венозная - в легкие и кожу, а смешанная - во все остальные органы тела. Таким образом, у земноводных нет полного разделения крови в желудочке, поэтому интенсивность жизненных процессов невысокая, а температура тела непостоянная.
Сердце у пресмыкающихся трехкамерное, однако полного смешения артериальной и венозной крови не происходит из-за наличия в нем неполной продольной перегородки. Отходящие от разных частей желудочка три сосуда - легочная артерия, левая и правая дуги аорты - несут венозную кровь к легким, артериальную - к голове и передним конечностям, а к остальным частям - смешанную с преобладанием артериальной. Такое кровообеспечение, а также малая способность к терморегуляции приводят к тому, что
температура тела пресмыкающихся зависит от температурных условий окружающей среды.
Высокий уровень жизнедеятельности птиц обусловлен более совершенной системой кровообращения по сравнению с животными предыдущих классов. У них произошло полное разделение артериального и венозного потоков крови. Это связано с тем, что сердце птиц четырехкамерное и полностью разделено на левую - артериальную, и правую - венозную, части. Дуга аорты только одна (правая) и отходит от левого желудочка. В ней течет чистая артериальная кровь, снабжающая все ткани и органы тела. От правого желудочка отходит легочная артерия, несущая в легкие венозную кровь. Кровь быстро движется по сосудам, газообмен происходит интенсивно, выделяется много тепла. Кровеносная система млекопитающих принципиальных отличий от таковой у птиц не имеет, В отличие от птиц, у млекопитающих от левого желудочка отходит левая дуга аорты.
Вопрос 22
РАЗВИТИЕ АРТЕРИАЛЬНЫХ ДУГ
Артериальные дуги, дуги аорты, кровеносные сосуды, закладывающиеся у зародышей позвоночных в виде 6-7 (у круглоротых до 15) парных боковых стволов, отходящих от брюшной аорты. А. д. проходят по межжаберным перегородкам на спинную сторону глотки и, сливаясь, образуют спинную аорту. Первые 2 пары артериальных дуг обычно рано редуцируются, у рыб и личинок земноводных они сохраняются в виде небольших сосудов. Остальные 4-5 пар артериальных дуг становятся жаберными сосудами. У наземных позвоночных из третьей пары артериальных дуг образуются сонные артерии, из шестой - лёгочные. У хвостатых земноводных обычно 4 и 5-я пары артериальных дуг образуют стволы или корни аорты, сливающиеся в спинную аорту. У бесхвостых земноводных и пресмыкающихся дуги аорты возникают только из 4-й пары артериальных дуг, а 5-я редуцируется. У птиц и млекопитающих редуцированы 5-я и половина 4-й артериальных дуг, у птиц аортой становится её правая половина, у млекопитающих - левая. Иногда у взрослых особей сохраняются зародышевые сосуды, соединяющие дуги аорты с сонными (сонные протоки) или с лёгочными (боталловы протоки) артериями.
Вопрос 23
Дыхательная система.
Большинство животных – аэробы. Диффузия газов из атмосферы посредством водного раствора осуществляется при дыхании. Элементы кожного и водного дыхания сохраняются даже в высших позвоночных животных. В ходе эволюции у животных возникли разнообразные дыхательные приспособления – производные кожи и пищеварительной трубки. Жабры и легкие – производные глотки.
ФИЛОГЕНЕЗ ОРГАНОВ ДЫХАНИЯ
Органы дыхания -жабры - расположены на верхней стороне четырех жаберных дуг в виде ярко-красных лепестков. Вода попадает в рот рыбы, процеживается через жаберные щели, омывая жабры, и выводится наружу из-под жаберной крышки. Газообмен осуществляется в многочисленных жаберных капиллярах, кровь в которых течет навстречу омывающей жабры воде.
Дышат лягушки легкими и кожей. Легкие представляют собой парные полые мешки с ячеистой внутренней поверхностью, пронизанной сетью кровеносных капилляров, где и происходит газообмен. Механизм дыхания у земноводных несовершенен, нагнетательного типа. Животное набирает воздух в ротоглоточную полость, для чего опускает дно ротовой полости и открывает ноздри. Затем ноздри закрываются клапанами, дно ротовой полости поднимается, и воздух нагнетается в легкие. Удаление воздуха из легких происходит благодаря сокращению грудных мышц. Поверхность легких у земноводных невелика, меньше поверхности кожи.
Органы дыхания -легкие(рептилии). Их стенки имеют ячеистое строение, что в значительной степени.увеличивает поверхность. Кожное дыхание отсутствует. Вентиляция легких более интенсивная, чем у земноводных, и связана с изменением объема грудной клетки. Дыхательные пути -трахея, бронхи - защищают легкие от иссушающего и охлаждающего воздействия воздуха, поступающего извне.
Легкие птиц представляют собой плотные губчатые тела. Бронхи, войдя в легкие, сильно в них ветвятся до тончайших, слепо замкнутых бронхиол, опутанных сетью капилляров, где
и происходит газообмен. Часть крупных бронхов, не разветвляясь, выходит за пределы легких и расширяется в огромные тонкостенные воздушные мешки, объем которых во много раз превосходит объем легких (рис. 11.23). Воздушные мешки расположены между различными внутренними органами, а их ответвления проходят между мышцами, под кожу и в полости костей.
Дышат млекопитающие легкими, которые имеют альвеолярную структуру, благодаря которой дыхательная поверхность превосходит поверхность тела в 50 раз и более. Механизм дыхания обусловлен изменением объема грудной клетки за счет движения ребер и свойственной млекопитающим особой мышцы - диафрагмы.
Вопрос 24
ФИЛОГЕНЕЗ ГОЛОВНОГО МОЗГА
Центральная нервная система рыб состоит из головного и спинного мозга. Головной мозг у рыб, как у всех позвоночных, представлен пятью отделами: передним, промежуточным, средним, мозжечком и продолговатым мозгом. От переднего мозга отходят хорошо развитые обонятельные доли. Наибольшего развития достигает средний мозг, осуществляющий анализ зрительных восприятий, а также мозжечок, регулирующий координацию движений и сохранение равновесии
Головной мозг земноводных имеет те же пять отделов, что и мозг рыб. Однако отличается от него большим развитием переднего мозга, который у земноводных разделен на два полушария. Мозжечок недоразвит в связи с малой подвижностью и однооб. разным характером движений земноводных.
Головной мозг пресмыкающихся по сравнению с таковым земноводных обладает лучше развитыми мозжечком и большими полушариями переднего мозга, поверхность которого имеет зачатки коры. Это обусловливает разнообразные и более сложные формы приспособительного поведения.
Головной мозг птиц отличается от мозга пресмьгкающихся большими размерами полушарий переднего мозга и мозжечка.
Головной мозг млекопитающих имеет относительно крупные размеры из-за увеличения объема полушарий переднего мозга и мозжечка. Развитие переднего мозга происходит за счет разрастания его крыши - мозгового свода, или коры мозга.
Вопрос 25
ФИЛОГЕНЕЗ ВЫДЕЛИТЕЛЬНОЙ И ПОЛОВОЙ СИСТЕМ
Органами выделения рыб служат парные лентовидные туловищные почки, расположенные в полости тела под позвоночником. Они утратили связь с полостью тела и удаляют вредные продукты жизнедеятельности, отфильтровывая их из крови. У пресноводных рыб конечным продуктом белкового обмена является ядовитый аммиак. Он растворяется большим количеством воды, и поэтому рыбы выделяют много жидкой мочи. Выведенная с мочой вода легко, восполняется за счет ее постоянного поступления через кожу, жабры и с пищей. У морских рыб конечным продуктом азотистого обмена служит менее ядовитая мочевина, выведение которой требует меньшего количества воды. Образовавшаяся в почках моча по парным мочеточникам оттекает в мочевой пузырь, откуда выводится наружу через выделительное отверстие. Парные половые железы - яичники и семенники - имеют выводящие протоки. Оплодотворение у большинства рыб наружное и происходит в воде
Органы выделения земноводных, как и у рыб, представлены туловищными почками. Однако в отличие от рыб они имеют вид уплощенных компактных тел, лежащих по бокам
крестцового позвонка. В почках имеются клубочки, которые отфильтровывают из крови вредные продукты распада (в основном мочевину) и одновременно важные для организма вещества (сахара, витамины и др.). Во время стока по почечным канальцам полезные организму вещества всасываются обратно в кровь, а моча поступает по двум мочеточникам в клоаку и оттуда в мочевой пузырь. После наполнения мочевого пузыря его мышечные стенки сокращаются, моча выводится в клоаку и выбрасывается наружу. Потери воды из организма земноводных с мочой, так же как и у рыб, восполняются поступлением ее через кожу. Половые железы парные. Парные яйцеводы впадают в клоаку, а семявыводящие каналы - в мочеточники.
Органы пресмыкающихся выделения представлены тазовыми почками, в которых общая фильтрационная площадь клубочков небольшая, в то время как протяженность канальцев значительная. Это способствует интенсивному обратному всасыванию отфильтрованной клубочками воды в капилляры крови. Следовательно, выделение продуктов жизнедеятельности у пресмыкающихся происходит с минимальными потерями воды. У них, как и у наземных членистоногих, конечным продуктом выделения является мочевая кислота, требующая для выведения из организма небольшого количества воды. Моча по мочеточникам собирается в клоаку, а из нее в мочевой пузырь, из которого в виде взвеси мелких кристаллов выводится наружу.
Выделение млекопитающих. Тазовые почки млекопитающих сходны по строению таковыми птиц. Моча с большим содержанием мочевины оттекает от почек по мочеточникам в мочевой пузырь, а из него выходит наружу.
Вопрос 26
Филогенез покровов тела:
Основные направления эволюции покровов хордовых:
1) дифференцировка на два слоя: наружный - эпидермис, внутренний - дерму и увеличение толщины дермы;
1) от однослойного эпидермиса к многослойному;
2) дифференцировка дермы на 2 слоя - сосочковый и сетчатый:
3) появление подкожно-жировой клетчатки и совершенствование механизмов терморегуляции;
4) от одноклеточных желез к многоклеточным;
5) дифференцировка различных производных кожи.
У низших хордовых (ланцетник) эпидермис однослойный, цилиндрический, имеет железистые клетки, выделяющие слизь. Дерма (кориум) представлена тонким слоем неоформленной соединительной ткани.
У низших позвоночных эпидермис становится многослойным. Нижний его слой - ростковый (базальный), клетки его делятся и восполняют клетки вышележащих слоев. Дерма имеет правильно расположенные волокна, сосуды и нервы.
Производными кожи являются: одноклеточные (у круглоротых рыб) и многоклеточные (у земноводных) слизистые железы; чешуя: а) плакоидная у хрящевых рыб, в развитии которой принимают участие эпидермис и дерма; б) костная у костных рыб, которая развивается за счет дермы.
Плакоидная чешуя снаружи покрыта слоем эмали (эктодермального происхождения), под которым находятся дентин и пульпа (мезодермального происхождения). Чешуя и слизь выполняют защитную функцию.
У земноводных кожа тонкая гладкая, без чешуи. Кожа содержит большое количество многоклеточных слизистых желез, секрет которых увлажняет покровы и обладает бактерицидными свойствами. Кожа принимает участие в газообмене.
У высших позвоночных в связи с выходом на сушу эпидермис становится сухим, имеет роговой слой.
У рептилий развиваются роговые чешуи, отсутствуют кожные железы.
У млекопитающих: хорошо развиты эпидермис и дерма, появляется подкожно-жировая клетчатка.
Вопрос 27
ФИЛОГЕНЕЗ ПИЩЕВАРИТЕЛЬНОЙ СИСТЕМЫ.
Питаются рыбы разнообразной пищей. Пищевая специализация отражается на строении органов пищеварения. Рот ведет в ротовую полость, в которой обычно имеются многочисленные зубы, расположенные на челюстных, небных и других костях. Слюнные железы отсутствуют. Из ротовой полости пища проходит в глотку, прободенную жаберными щелями, и по пищеводу попадает в желудок, железы которого обильно выделяют пищеварительные соки. У некоторых рыб (карповые и ряд других) желудка нет и пища поступает сразу в тонкий кишечник, где под влиянием комплекса ферментов, выделяемых железами самого кишечника, печени и поджелудочной железы, происходит расщепление пищи и всасывание растворенных питательных веществ. Дифференцировка пищеварительной системы земноводных осталась примерно на том же уровне, что и у их предков - рыб. Общая ротоглоточная полость переходит в короткий пищевод, за ним расположен слабо обособленный желудок, переходящий без резкой границы в кишечник. Кишечник заканчивается прямой кишкой, переходящей в клоаку. Протоки пищеварительных желез - печени и поджелудочной железы - впадают в двенадцатиперстную кишку. В ротоглоточную полость открываются протоки отсутствующих у рыб слюнных желез, смачивающих ротовую полость и пищу. С наземным образом жизни связано появление в ротовой полости настоящего языка - основного органа добычи пищи.
В пищеварительной системе пресмыкающихся лучше, чему земноводных, выражена дифференцировка на отделы. Захват пищи производится челюстями, имеющими для удержания добычи зубы. Ротовая полость лучше, чем у земноводных, отграничена от глотки. На дне ротовой полости расположен подвижный, раздвоенный на конце язык. Пища смачивается слюной, что облегчает ее заглатывание. Пищевод в связи с развитием шеи длинный. Отграниченный от пищевода желудок имеет мускулистые стенки. На границе тонкой и толстой кишок имеется слепая кишка. Протоки печени и поджелудочной
железы открываются в двенадцатиперстную кишку. Время переваривания пищи зависит от температуры тела пресмыкающихся.
Пищеварительная система млекомитающихся. Зубы сидят в ячейках челюстных костей и подразделяются на резцы, клыки и коренные. Ротовое отверстие окружено мясистыми губами, что свойственно только млекопитающим в связи со вскармливанием молоком. В ротовой полости пища, кроме прожевывания зубами, подвергается химическому воздействию ферментов слюны, а затем последовательно переходит в пищевод и желудок. Желудок у млекопитающих хорошо обособлен от других отделов пищеварительного тракта и снабжен пищеварительными железами. У большинства видов млекопитающих желудок разделен на большее или меньшее число отделов. Наиболее сложен он у жвачных парнокопытных. Кишечник имеет тонкий и толстый отделы. На границе тонкого и толстого отделов отходит слепая кишка, в которой происходит сбраживание клетчатки. Протоки печени и поджелудочной железы открываются в полость двенадцатиперстной кишки.
Вопрос 28
Эндокринная система.
В любом организме вырабатываются соединения, разносящиеся по всему организму, имеющие интегративную роль. У растений есть фитогормоны, контролирующие рост, развитие плодов, цветов, развитие пазушных почек, деление камбия и др. Фитогормоны есть у одноклеточных водорослей.
Гормоны появились у многоклеточных организмов, когда возникли специальные эндокринные клетки. Однако химические соединения, играющие роль гормонов, были и раньше. Тироксин, трийодтиронин (щитовидная железа) обнаружены у цианобактерий. Гормональная регуляция у насекомых изучена плохо.
В 1965 году Вильсон выделил инсулин из морской звезды.
Оказалось, что дать определение гормону очень трудно.
Гормон – это специфическое химическое вещество, выделяемое особыми клетками в определенном участке тела, которое поступает в кровь и затем оказывает специфическое действие на определенные клетки или органы-мишени, расположенные в других областях тела, что приводит к координации функций всего организма в целом.
Известно большое количество гормонов млекопитающих. Они делятся на 3 основные группы.
Феромоны. Выделяются во внешнюю среду. С их помощью животные принимают и передают информацию. У человека запах 14 - окситететрадекановой кислоты четко различают только женщины, достигшие половой зрелости.
Наиболее просто организованные многоклеточные организмы – например, губки тоже имеют подобие эндокринной системы. Губки состоят из 2 слоев – энтодермой и экзодермой, между ними располагается мезенхима, в которой содержатся макромолекулярные соединения, характерные для соединительной ткани более высокоорганизованных организмов. В мезенхиме есть мигрирующие клетки, некоторые клетки способны секретировать серотонии, ацетилхолин. Нервная система у губок отсутствует. Вещества, синтезируемые в мезенхиме, служат для связи отдельных частей организма. Координация осуществляется за счет перемещения клеток по мезенхиме. Есть также и перенос веществ между клетками. Заложена основа химической сигнализации, которая характерна для остальных животных. Самостоятельных эндокринных клеток нет.
У кишечнополостных имеется примитивная нервная система. Первоначально нервные клетки выполняли нейросекреторную функцию. Трофическую функцию, осуществляли контроль роста, развития организма. Затем нервные клетки стали вытягиваться и образовали длинные отростки. Секрет выделялся около органа-мишени, без переноса (т.к. не было крови). Эндокринный механизм возник раньше проводникового. Нервные клетки были эндокринными, а потом получили и проводниковые свойства. Нейросекреторные клетки был первыми секреторными клетками.
Первичноротые и вторичноротые вырабатывают одинаковые стероидные и пептидные гормоны. Принято считать, что в процессе эволюции из одних полипептидных гормонов могут возникнуть новые (мутации, дупликации генов). Дупликации менее подавляются естественным отбором, чем мутации. Многие гормоны могу синтезироваться не в одной железе, а в нескольких. Например, инсулин вырабатывается в поджелудочной железе, подчелюстной железе, 12-перстной кишке и других органах. Существует зависимость генов, контролирующих синтез гормонов от положения.
